Костная система
За последние десятилетия ведомости структурно функциональные особенности костной системы высших позвоночных значительно дополнились новыми исследованиями. Современное значение костной системы – не только опора и рычаги передвижение, но и орган трофической, кроветворной функции, электролитического баланса, центральный орган иммунной системы которые генетически возникли в связи с движением и существуют для обеспечения двигательной активности животного и сохранения внутреннего гомеостаза и жизни всего организма.
Костная система, стала одной из интегрирующих ситем, предопределяет морфофункциональное состояние всего организма [332]. Она обеспечивает благополучие организма. Это объясняется тем, что она полифункциональная и, выполняя наиболее присущую ей древнюю функцию – биомеханическую, обеспечивает иммунную защиту организма, являясь универсальным органом гемоиммунопоэза.
Костные органы, несмотря на их разъединенность, являются единственными целыми, что определяется особенностью строения их системы венозных сосудов. Экстраосальные венозные сосуды обеспечивают общность кровотока не только костной системы, но и всех органов и тканей всего организма. Доказательством этого явления является возможность наполнения контрастной жидкостью всю венозную систему сосудов организма при условии ее введения через любую часть костного органа, которая содержит красный костный мозг [133, 81].
Внутрикостные кровеносные сосуды, которые составляют 50% массы костного органа, обеспечивают необходимое микроокружение для образования и развития клеточных структур эритроидного и лимфоидного ряда. Характерно, что в костных органах обнаружены синусоидные капилляры, стенка которых построена из высокого эндотелия, что дало возможность исследователям определить их как венулы с высоким эндотелием. Только у них проходят все интимные процессы разного рода образования и трансформации стволовых полипотентных клеток, которые обеспечивают начало иммунокомпетентности структур организма [81, 133].
Рост и развитие костных органов проходит непрерывно в течение всей жизни животного соответственно действию сил сжатия и растяжения, которые возникают в период двух фаз работы аппарата движения, – динамической и статической [239, 252]. Изменяется действие биомеханической нагрузки на костный орган – изменяется ее архитектоника, как и всей костной системы. Отсутствие действия биомеханической нагрузки на костный орган, что связанная с движением животного, приводит, прежде всего, к потере его твердости – в результате деминерализации.
Усиление двигательной активности животного, соответственно генетическим запросам, напротив, приводит к оптимизации всех его функций. Сущность процессов, которые происходят в костных органах заключается в их постоянном разрушении и возобновлении (резорбции и регенерации), то есть в разрушении старых структур и трансформации в новые, которые отвечают условиям действующих биомеханических нагрузок. Достояние в филогенезе свойств постоянного разрушения и возобновления обеспечили не только прочность и легкость костных органов, но и, одновременно, привели к участию костной системы в обмене веществ, гемоиммунопоезе, гомеостазе и частично в крово – лимфообращении и импульсации мозга.
Следовательно, биомеханические нагрузки, которые действуют на костную систему ее костные органы, в частности, являются тем необходимым фактором и стимулятором, которые кроме основной, филогенетически наиболее древней функции движения определяют в ней кроветворную, трофическую и функции обменную веществ и иммунной защиты.
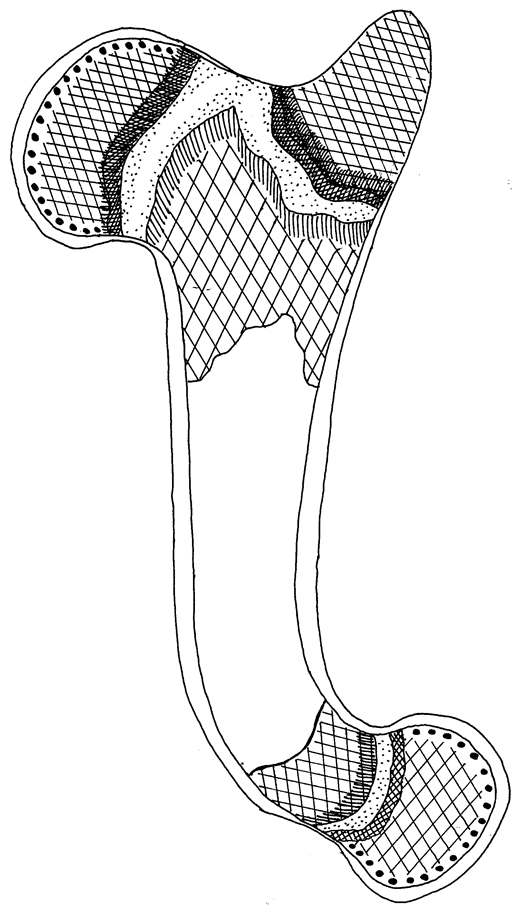
Костный орган насколько полифункциональный, настолько и полиморфологичный, который включает в себя образование разных по происхождению и физиологическим назначением компонентов: собственно костная ткань (компактная и губчатая), костный мозг (остеобластичний, красный и желтый), надкостница, эндост, гиалиновые хрящи (суставные и метафизарные), кровеносные сосуды и нервы (рис. 101).

|
![]()
![]()
![]()
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
||
|
Рис. 101. Строение трубчатого костного органа (Криштофорова, 1987)
1-суставной хрящ, 2-метафизарный хрящ, 3- проксимальный эпифиз, 4- дистальный эпифиз, 5-апофиз, 6- диафиз, 7- диаметафизарная субхондральная кость, 8- эпитаметафизарная субхондральная кость, 9- субхондральная кость суставного хряща, 10- костномозговой участок диафиза.
Костная ткань является разновидностью соединительной ткани с характерной особенностью межклеточных структур. Они образованы биополимерными макромолекулами тропоколагена, которые соединены с кристаллами гидрооксиапатита. До пяти молекул тропоколагена совмещаются между собой спиралевидно и соединяются гидрооксиапатитом.
Гликозаминогликаны, гликопротеины и протеогликаны образуют фибриллярную вещество, соединяя между собой фибриллы, и формируют костные пластинки, которые в свою очередь строят определенную систему, – остеон, что имеет очень сложное строение. Костные пластинки, при соединении между собой, могут образовывать не только остеон, а трабекули (перекладины), которые, как правило, расположенные в направлении действия силы притяжения. При этом образуется губчатая костная ткань. Губчатая костная ткань способна рассеивать и гасить напряжение, которое возникает при биомеханических нагрузках.
Более плотное размещение костных пластин (в виде остеонов), образует компактную костную ткань, которая размещается на поверхности костного органа и утолщается в местах, где максимально действуют нагрузки на сгибание, как это имеет место в длинных трубчатых костях.
Соотношение в костных органах компактной и губчатой костной ткани отражается на активности метаболизма. 1 г губчатой костной ткани имеет общую площадь активной поверхности 300 мм 2, компактной – только 80 мм2. В целом же общая масса губчатой костной ткани в организме млекопитающих в четыре разы меньше, по сравнению с компактной, однако совокупная поверхность первой в 9,4 разы большая чем второй. Если учесть, что у новорожденных млекопитающих компактная костная ткань имеет характерное сетчатое строение, то можно допустить, что она полностью способна обеспечить необходимые обменные процессы, которые наиболее интенсивно протекают в новорожденный период, предопределенные необходимостью морфофункционального становления органов и систем, и в первую очередь скелета для реализации генетических потребностей в локомоции.
Известно, что 30% белков организма и до 50% коллагена находится в костной системе (в коже – 40%, а в других органах – 10%). Коллаген составляет 90% всего органического матрикса костных органов. Из всех неорганических веществ 98% находится в костных органах (Ca – 99%, P – 87%, Mg– 58%), которые включают всю таблицу химических элементов Менделеева в определенных соотношениях.
Костные органы (костная система в целом) выполняют одну из сложных задач, рядом с органами дыхания и выделения, в поддержке кислотно-щелочного равновесия, поставляя фосфор и карбонаты в качестве буферов. В обезвоженном костном органе содержится кальция – 27%, фосфора – 12%, магния – 0,5%. В нем также оказываются не минерализованные пространства в лакунах и канальцах по периферии клеток, которые наполнены межклеточным веществом, которое наиболее активно принимает участие в обмене веществ.
Гистогенез, ремоделирование и перестройка костных органов обеспечивается анаболическими и катаболическими процессами, интенсивность которых преобладает друг над другом в определенных средах и при определенных уровнях жизнедеятельности их клеточных структур: полипотентных стволовых, миелоидных, остеобластов, остеоцитов, остеокластов, хондробластов, хондроцитов и других.
Остеогенные клетки содержатся в надкостнице (камбиальный слой), эндоста, на поверхности гаверсовых (костных) канальцев и по периферии перекладин первичной губчатой ткани. Названные клетки принимают участие в образовании костной ткани.
Остеобласты – клетки, которые синтезируют органический матрикс и предопределяют его минерализацию. Они способны образовывать волокна, которые содержат значительное количество фосфатазы, которая определяет их роль в минерализации костного матрикса. В плазматических мембранах остеобластов содержится Са+2 – зависимая АТФаза, детерминируя течение Са+2 в костных клетках. Этот фермент определяет их функцию. Однако для активности самого фермента необходимо присутствие Mg+2. Вот почему важно для выполнения функций и перестройки костных органов наличие определенного соотношения этих элементов.
Остеобласты синтезируют также проколаген, аморфное вещество и ряд ферментов. Проколаген превращается в коллаген и вследствие этого образуется остеоидная (не минерализованная) ткань способная к декальцинации.
Остеоцити – это зрелые остеобласты, которые заключены в матрикс. Остеоцити обеспечивают функцию органического матрикса и стабильность минерального компонента, а также принимают участие в регуляции определенного уровня Са+2 в крови.
Интенсивность процессов метаболизма также предопределяется течением межклеточного вещества между ними и костными канальцами.
Остеокласти – клетки, которые принимают участие в резорбции костной ткани, что необходимо для ее морфогенеза и постоянного обновления. Это многоядерные клетки, которые выделяют разрушающие ферменты и кислоты, способные к поглощению убломков костного матрикса и их последующего переваривания.
Хондробласти и хондроцити – клетки, которые являются производными мезенхимы, содержатся в толще метафизарных и суставных хрящей костных органов на разных этапах развития.
Условием для соответствующей полифункциональности костных органов является определенное взаимодействие всех клеточных структур. Образование костной ткани (остеогенез) очень сложный динамический процесс, который протекает в несколько фаз: 1. Формирование специфических клеточных популяций; 2. Образование клеточного матрикса; 3. Его минерализация. Весь процесс остеогенеза предопределяется целым рядом факторов и морфогенетических белков.
В костных органах новорожденных животных очень интенсивно происходит остеогенез. Превращение эндесмальной ткани в костную сопровождается образованием последней сетчатой структуры. Костные трабекули образуют ячейки, в которых содержится соединительная ткань, пронизанная кровеносными сосудами. В последующем, с возрастом животных, костные трабекули утолщаются, а ячейки, наоборот, уменьшаются. Между внешней и внутренними пластинами образуются толстые трабекули, ячейки увеличиваются, а соединительная ткань редуцируется, что приводит к образованию параназальных синусов. Таким образом, образуются кости черепа, за исключением клиновидной и затылочной. В этих органах, как других костных органах, происходит смешанный остеогенез. На первом этапе при условии образования костной манжеты или эпифизарных центров окостенения происходит энхондральный остеогенез. Хрящевая ткань интенсивно трансформируется в костную. При условии образования диафизарного центра окостенения в виде манжеты, от нее, в проксимальном и дистальном направлениях, хрящевая ткань разрушается, а губчатая костная ткань интенсивно образуется, однако ее трабекулы содержат значительное количество разрушительного хряща. Ячейки, которые образуются такими костными трабекулами, заполнены кровеносными сосудами, а также остеобластичним костным мозгом. В дальнейшем первичное губчатое вещество трансформируется во вторичное, что предопределяет замещение остеобластического костного мозга на красный. С образованием надкостницы происходит интенсивный эндесмальный остеогенез, который приводит к образованию компактной костной ткани. На первом этапе камбиальные клетки надкостницы трансформируются в остеобласты, которые продуцируют остеоид в виде прямых удлиненных образований. Между ними содержатся ячейки удлиненные и разной величины, заполненные соединительной тканью, среди которой оказываются сосуды разного калибра. Костные трабекули утолщаются, а ячейки постепенно уменьшаются в размерах. Таким образом, образуется компактная костная ткань, что наибольшего развития достигает в диафизах трубчатых костных органов конечностей суточных жеребят, телят, в меньшей степени поросят. В состоянии начального развития она находится у суточных щенков собак. У животных новорожденного периода компактная костная ткань диафиза трубчатых костных органов значительно утолщается. Промежутки между костными пластинками, формируя между ними щели.
На конец новорожденного периода компактная костная ткань образуется таким же образом по периферии тел позвонков, а также участков грудины. Постепенно компактная костная ткань, которая имеет сетчатое строение, пластинчатой, для которой характерно наличие остеонов. Следовательно, компактная костная ткань костных органов образуется при эндесмальном остеогенезе.
В период остеогенеза, костная ткань имеет разное строение. В ней различают зрелые и незрелые структуры. Незрелые (примитивные) - построены из ретикуло-фиброзной ткани (пучковой сетчато-волокнистой), которая наиболее лабильна и активна в метаболических процессах [259]. Ретикулофиброзная ткань содержит большое количество остеоцитов и коллагеновых волокон, размещенных без определенного порядка. В зрелых костных структурах остеоциты находятся, по своему размещению, в прямой зависимости от направления действия сил упругости и натяжения, образуют пластинки, которые образуют трубочки, – остеоны. Третий вид костной ткани переходный, имея строение какое-то среднее между незрелой и пластинчатой (смешанная костная ткань). У новорожденных продуктивных животных костные органы образованы ретикуло-фиброзной (незрелой) костной тканью, которая наиболее способна к ремоделяции.
Внешне костные органы покрыты надкостницей, через которую обеспечивается их кровоснабжение и иннервация. Внутренний слой надкостницы (остеогенний) обеспечивает утолщение костных органов в результате эндесмального остеогенеза. Оно также увеличивает их прочность, особенно в участках диафизов трубчатых костей конечностей [341]. Внешний слой надкостницы образован из плотной соединительной ткани.
Эндост содержится в середине костных органов, выстилая из внутреннюю поверхность компактной костной ткани и поверхность трабекул губчатой. Он образован из тонкого слоя рыхлой соединительной ткани, которая содержит значительное количество камбиальных клеток разных популяций. Вопросы о наличии эндоста и его функциях еще не достаточно исследованы и дискуссионны. Однако исследования свидетельствуют, что эндост, в определенной мере, активизирует гемопоетическую функцию костного мозга.
Хрящевая ткань и ее взаимодействие с костной, является одним из факторов поддержки кроветворной функции костного мозга. Она образует суставные и метафизарные хрящи, которые относятся к гиалиновым (стекловидным). В состав суставных и метафизарных хрящей входят клетки (хондробласти и хондроцити) и межклеточное вещество, которое состоит из основного вещества и коллагеновых волокон. Основное вещество хрящевой ткани содержит протеин-полисахаридные комплексы, которые превращают дисперсную фазу коллоида в жидкость, которая является водным раствором электролитов. Концентрация их практически отвечает наличию в диализатах плазмы крови и синовии. Сочетание дисперсной и водяной фаз в коллоидной системе обеспечивает лабильность основного вещества хрящей за счет возможностей перестройки полимерных цепей и изменения содержания воды. Коллагеновый компонент матрикса хрящей костных органов, формируя ретикулярные сетки, ограничивает свободное перемещение больших молекул и их комплексов, что осложняет обмен веществ не имеющих собственного кровоснабжения.
Суставной хрящ размещен между субхондральною костной тканью (СХКТ) и синовией суставной полости, тогда как метафизарный находится между двумя СХКТ – эпифизарной и диафизарной (сбоку эпифиза и диафиза). Возможно, это является основным фактором, который определяет разницу в функции этих хрящей. Одной из главных особенностей суставных и метафизарных хрящей у млекопитающих есть их морфологическая полярность.
Характерно, что еще у плодов продуктивных животных в суставном хряще выделяются зоны с особенностью структуры межклеточных элементов и основного вещества: поверхностная; камбиальная; зона эмбрионального хряща; зона гипертрофированных хондроцитов; гидратации и деструкции хондроцитов; промежуточная зона и зона СХКТ. С возрастом структура суставного хряща несколько изменяется. Уменьшаются его пролиферативные потенции (что приводит к его утончению), количество хондроцитов, исчезает камбиальная зона. В поверхностной зоне выделяется внешний безклеточный слой, который состоит из матрикса, который иногда содержит остатки хондроцитов.
Полярное строение суставного хряща определяется образованием сустава. Полярность метафизарного хряща формируется значительно позже (до конца молочного периода) и определяется структурой СХКТ.
Среди других важных особенностей имеет значение питание хрящей. В суставном хряще отсутствует граница со стороны суставной полости, потому он “открыт” для обмена веществ с синовиальной жидкостью. Проницательность суставного хряща для белков, ферментов и электролитов синовиальной жидкости обеспечивается степенью его "пористости" – архитектоникой собственных молекул и агрегатов между которыми могут проникать молекулы синовии.
Суставной хрящ и его СХКТ обеспечивают рост эпифизов костных органов. С момента превращения СХКТ суставного хряща в запирающую пластинку остеонного строения, рост эпифизов костных органов останавливается.
Метафизарный хрящ вместе с диаметафизарной СХКТ образуют ростковую зону, которая обеспечивает интенсивный рост диафиза в длину. В ней определяется наиболее развитое микроциркуляторное русло, которое обеспечивает интенсивное образование ретикуло-фиброзной костной ткани с последующей импрегнацией минеральными веществами [76].
Структурные изменения суставных и метафизарных хрящей связанны с возрастом и активным дваижением животного. Ее уменьшение (гиподинамия) предопределяет интенсивное утончение суставных хрящей и полное разрушение метафизарных, что приводит к синостозу [135]. Необходимо отметить, что синостоз между диафизом и эпифизами трубчатых костей конечностей уже наблюдается у суточных домашних животных.
Синостоз имеет свои особенности. Он образуется одиночными сосудами, которые проходят сквозь толщу метафизарного хряща. На поверхности хряща, формируются тонкие пластинки остеида, что размещается вокруг проникающих кровеносных сосудов. Таких тонких иглообразных образований встречается 5-6 на гистологическом срезе. С возрастом телят количество иглообразных синостозов растет, а пластинки остеида утолщаются. Наличие таких одиночных синостозных образований не препятствует росту костных органов в длину, о чем свидетельствует увеличение их параметров. Однако в настоящее время не известно, чем предопределен такой синостоз костных органов. Можно допустить, что он связан с увеличением кровоснабжения при образовании эпифизарных центров окостенения.
У взрослых животных синостоз эпифизов с диафизами костных органов достаточно длительный во времени и проходит асинхронный. Интенсивность роста и формирования хрящей также предопределена интенсивностью образования их СХКТ, что приводит к уменьшению микроциркуляции и снижению метаболизма.
Необходимо отметить обнаруженную закономерность длительного становления полифункциональности костных органов, всей костной системы, которая дает возможность понять ее важное значение для существования организма.
В филогенезе костная система формировалась под действием сил гравитации и двигательной активности животных, выполняя, главным образом, биомеханическую функцию. Костные органы у наземных высших позвоночных животных обогащаются новым структурным элементом – костным мозгом, который в начале является остеобластическим в связи с появлением скелета в гравитационном положении, обеспечивая более интенсивные процессы ремоделяции, а затем становится кроветворным ибо первым отвечает на изменения физической нагрузки при движении – органом универсального гемоиммунопоэза, становится уже красным костным мозгом, заменяя печень. Необходимо отметить, что костный мозг, который является неотъемлемой частью костного органа, становится одной из главных звеньев, которые объединяют организм в единое целое с окружающей средой, выполняя функцию кроветворения и иммуногенеза. На какое либо изменение внутренней или внешней среды костный мозг реагирует изменением морфологического состава крови и морфофункционального статуса лимфоидных органов. Кроме этого, само присутствие костного мозга значительно усиливает упругость и прочность костных органов и их регенерацию, в связи с чем красный костный мозг может стать желтым костным мозгом.
Выделяют три вида костного мозга, которые постепенно возникали в филогенезе [33, 133, 134, 135, 157, 171]. Остеобластичний костный мозг возникает у наземных высших позвоночных животных и выполняет костеобразующую функцию. Его наибольшее количество находится в костных органах плодов и новорожденных животных. С выходом животных в наземную среду, в условия действия земного притяжения, остеобластический костный мозг трансформируется в красный (кроветворный). У плодов, новорожденных птиц и млекопитающих все костные органы содержат красный костный мозг. Только с возрастом млекопитающих, в конце молочного периода, в диафизарный части трубчатых костей красный костный мозг замещается желтым (жировым), функция которого в настоящее время не установлена (см. раздел 3.1).
У взрослых птиц и млекопитающих костный мозг составляет 4-5% живой массы или 40-50% массы костной системы. Однако определено, что чем больше подвижная активность животного, тем больше костного мозга содержится в костной системе (П. А. Коржуев, 1961, 1968).
Внешняя форма и структура костных органов отвечает биомеханическим нагрузкам, которые действуют на отдельную часть или на весь организм. Точно эти нагрузки, связанные всегда с движением животного (его динамикой и статикой) и обеспечивают костным органам полифункциональность. Деформирующие силы, которые действуют на костный орган, образуются под действием живой массы и мышцами, которые крепятся на нем. В зависимости от этого изменяются морфологические, биофизические и биохимические свойства костных органов приобретая свои многочисленные функции.
Упругие деформации костных органов и их отдельных частей разные: в губчатой костной ткани они наибольшие (в 7 раз больше чем в компактном), что предопределяется максимальным количеством костного мозга в ячейках губчатой костной ткани кровеносных сосудов и нервов, оптимальными условиями питания, кроветворения и иммуногенеза и взаимоувязанных с двигательной активностью животных. С этой точки зрения, у неонатальных животных биологические закономерности становления костей, как органов, полностью зависят от биомеханических нагрузок связанных с движением [152, 333, 334].
Необходимо отметить, что относительная масса костной системы у новорожденных продуктивных животных значительно больше по сравнению с взрослыми (табл. 137, 138).
Таблица 137
Живая масса и масса костных органов бычков черно-рябой породы (Б. В. Криштофорова, 1987)
|
Возраст животных, суток |
Живая |
Абсолютная масса костных органов, кг |
Относительная масса костных органов % |
|
Новорожденные |
32,50±1,26 |
6,34±0,81 |
19,50 |
|
30 |
46,00-47,33 |
8,76-8,91 |
18,82-19,04 |
Таблица 138
Масса костных органов телочек красной степной породы (П. Н. Гаврилин, 1992)
|
Возраст, суток |
Абсолютная |
Относительная |
Суточный |
Относительный |
|
Новорожденные |
7,25±0,63 |
21,05 |
– |
– |
|
10 |
8,22-8,50 |
18,48-20,23 |
97,00-150,00 |
13,37±21,37 |
Она составляет 19-23% у телят, поросят, ягнят и цыплят и несколько больше у жеребят – (30-33%), тогда как у взрослых – 8-10%. Жизнеспособность неонатальных производительных животных полностью зависит от интенсивности остеогенеза во внутриутробный период онтогенеза.
Интенсивность замещения хрящевой ткани костной определяет пренатальное развитие всего организма на определенном этапе развития. У 2-х месячного эмбриона хрящевой скелет начинает замещаться костным. И именно с этого времени в костных органах, особенно конечностей, появляется красный костный мозг. Установлено, что одной из причин высокого заболевания новорожденных животных есть нарушение пренатального остеогенеза, который предопределяет задержку функции кроветворения и иммуногенеза.
У новорожденных продуктивных животных костные органы, рядом с биомеханической выполняют функции кроветворения, иммуногенеза, и активно участвуют в обмене веществ. Однако, реализация генетических потребностей в локомоции неонатальных животных, особенно матуронатних, является основным фактором, который обеспечивает не только интенсивность кровообращения, обмен энергии, импульсацию мозга, но и перестройку самих костных органов (трансформацию утробных структур на структуры в соответствии с генетической детерминацией неонатального периода).
Костная система, отдельные костные органы новорожденных животных значительно отличаются от таких взрослыми, не только внешне, но и внутренним строением [52-59, 65]. У новорожденных телят костные органы построены из ретикулофиброзной костной ткани. Компактная костная ткань имеет выраженное сетчатое строение (рис. 102). Костные балки анастомозують между собой. В их толще выделяются молодые остеоцити округлой формы с отростками на поверхности цитоплазмы. Между костными балками содержится рыхлая волокнистая соединительная ткань, в которой проходят кровеносные сосуды и нервы. Компактная костная ткань больше всего развитая в средней трети диафиза трубчатых костных органов конечностей.
Губчатая костная ткань содержится в эпифизах, проксимальных и дистальных частях диафиза трубчатых костей, и во всех костных органах осевого скелета. Характерной особенностью губчатой костной ткани костных органов новорожденных животных является то, что в них содержится значительное количество разрушительной хрящевой ткани.
В трабекулах губчатой костной ткани находится значительное количество хрящевой ткани. В трубчатых костях конечностей ее количество достигает больше 15%. Особенно ее количество растет в диаметафизарной субхондральной кости (рис. 103).
Характерно, что масса отдельных частей скелета у новорожденных животных неодинакова. У суточных телят осевой скелет составляет 38,7%, а скелет конечностей – 61,60% от общей массы скелета.
Отношение абсолютной массы позвоночного столба составляет 16,6%, скелета головы достигает 13,4%, ребер и грудины – 9,4% от массы всего скелета. Масса осевого скелета у новорожденных телят значительно меньше (8,90%) массы скелета конечностей (12,47%) (табл. 139, 140).
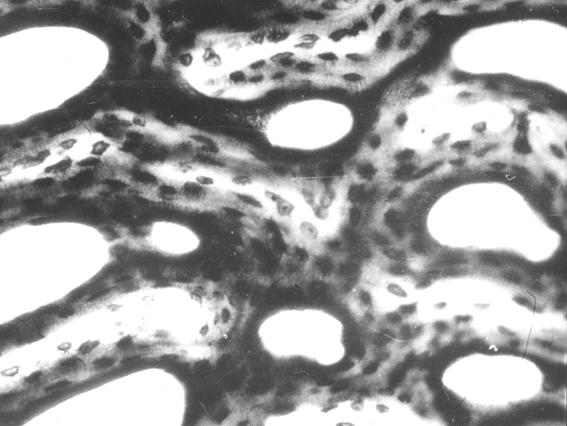
Рис. 102. Гистотопограмма компактной костной ткани теленка (сутки). Импрегнация серебром. МБИ-6, х400 (Б. В. Криштофорова, 1987).

Рис. 103. Гистотопограмма метафизарного хряща и субхондральной кости плечевой кости теленка (сутки). Гематоксилин и эозин. МБИ-6, х400 (Б. В. Криштофорова, 1987).
Таблица 139
Масса осевого и скелета конечностей телочек красной степной породы (П. Н. Гаврилин, 1992)
|
Возраст |
Осевой скелет |
Скелет конечностей |
Осевой скелет |
Скелет конечностей |
|
Абсолютная масса, кг |
Относительная масса % |
|||
|
Новорожденные |
2,81±0,29 |
4,44±0,40 |
8,16 |
12,39 |
|
10 суточные |
3,51–4,04 |
4,70±4,76 |
7,91–9,33 |
10,58–10,98 |
|
Суточный прирост, г |
Относительный прирост % |
|||
|
Новорожденные |
– |
– |
– |
– |
|
10 суточные |
70,00–123,00 |
26,00–32,00 |
24,91–43,77 |
5,85–7,20 |
Таблица 140
Масса скелета грудных и тазовых конечностей телочек красной степной породы (П. Н. Гаврилин, 1992)
|
Возраст |
Грудные |
Тазовые |
Грудные |
Тазовые |
|
А. Абсолютна масса, кг |
||||
|
Новорожденные |
1,85±0,16 |
2,85±0,24 |
5,39 |
7,49 |
|
10 суточные |
1,87–1,96 |
2,79–2,83 |
4,21–4,54 |
6,36–6,44 |
|
Б. Суточный прирост, г |
||||
|
Новорожденные |
– |
– |
– |
– |
|
10 суточные |
2,00–11,00 |
21,00–25,00 |
1,08–5,94 |
8,13–9,67 |
Относительная сама черепа (к осевому скелету) составляет 13%. Скелет головы новорожденных телят имеет значительную разницу, по сравнению с взрослыми. Длина от лобного гребня до подбородочного отверстия составляет 26,8%, высота скулового гребня – 21,3%, расстояние от середины края нижней орбиты к роговым отросткам – 21,3% длине взрослой коровы. Затылочная, клиновидная и решетчатая кости содержат хрящевую ткань, кроветворные островки в губчатой костной ткани. Между телами затылочных и клинообразной костей содержится слой хрящевой ткани. Остальные кости черепа отделяются одна от другой тонкими соединительнотканными прослойками. Гребни, бугры, линии к которым крепятся мышцы, слабо выраженные. Носовые пазухи незначительны, за исключением верхнечелюстной, что придает кругообразную форму мозговому скелету головы.
Шейный отдел достигает 5,62% массы осевого скелету суточных телят. Его абсолютная масса отвечает таким грудным позвонкам. Максимальную абсолютную массу имеет эпистрофей, меньшую четвертый, пятый и шестой позвонки, а минимальную – седьмой. Атлант имеет четыре центра окостенения – по два в каждой дужке, которые разделены прослойками хряща. В следующих позвонках – три центра окостенения (на теле и два на дужке). В позвонках головки и ямки покрыты гиалиновым хрящом, который переходит в волокнистый, по центру которого содержится пульпозное ядро. На гистотологичких срезах наличие волокнистого хряща свидетельствует о границе между телами позвонков. Головки и ямки позвонков вдоль всего позвоночного столба не выражены (плоские), что предоставляет последнему значительную подвижность (дорзомобильность).
В грудном отделе осевого скелета ребра очень подвижные, соединяются с позвонками. Их масса отвечает массе грудных позвонков. Наиболее развитое первое ребро и менее всего – последнее. Головка и бугорок ребра содержат хрящевую ткань (рис. 104). Привлекает внимание последнее ребро. Оно достаточно короткое и его вентральный конец размещается между мышцами брюшной стенки, но пальпируется легко. Степень его пренатального остеогенеза учитывается при определении морфофункционального состояния новорожденных, особенно телят (рис. 105, 106).

Рис. 104. Рентгенограммы 1, 2, 7, 10 ребер суточной телочки (П. Н. Гаврилин, 1992): 1 – первое ребро; 2-7 ребро; 3-13 ребро.
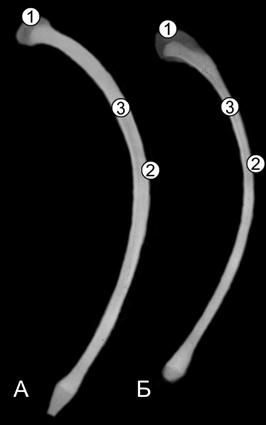
Рис. 105. Рентгенограммы последних костных ребер телочек (сутки) пренатальной развитой (А) и пренатальной недоразвитой (Б) групп (Ж. Г. Грабчак, 2004): 1 – хрящевые головка и бугорок; 2 – компактная костная ткань; 3 – губчатая костная ткань.
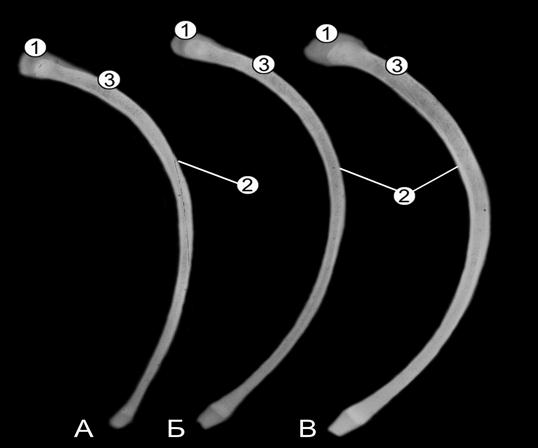
Рис. 106. Рентгенограммы последних костных ребер 5- (А), 10- (Б) и 20- (В) телочек (сутки) (Ж. Г. Грабчак, 2004): 1 – хрящевая головка и бугорок; 2 – компактная костная ткань; 3 – губчатая костная ткань.
У пренотальных недоразвитых суточных телят и поросят последнее ребро имеет значительно меньшую длину. Позвоночный конец образован хрящевой тканью. С возрастом животных, только у 10-суточных телят в позвоночной части последнего ребра появляется губчатая костная ткань. Однако головка и бугорок ребра еще образованы хрящевой тканью. Вентральная же часть ребра утолщена. Красный костный мозг размещается в средней части ребра, а остеобластичний в позвоночной и частично вентральной.
Стернальные ребра соединяются с грудиной. Грудина имеет значительно более тонкое тело по сравнению с взрослыми животными. Ее части разделены значительными по толщине и широкими прослойками хрящевой ткани (рис. 107, 108). Толщина прослоек между частями грудины больше у недоразвитых суточных телят. С возрастом животных части грудины увеличиваются, а прослойки хрящевой ткани истончаются. У пренатальных недоразвитых поросят на рентгенограммах, находятся части грудины разделенные на две, что свидетельствует о парных центрах их окостенения, которые сливаются в одно целое [287]. Размещение костного мозга (до 99%) подобно позвонкам и ребрам. Хрящевые прослойки частей груднини представлены гиалиновой хрящевой тканью. Вместе с СХКТ они образуют ростковые зоны костного органа. В центре прослойки находится основная зона хряща. Грудные позвонки, ребра и грудина образуют грудную клетку слегка приплюснутую по бокам и напоминающую по форме таковую хищников. Краниально она значительно уже, чем каудально, изгиб ребер незначителен.
Поясничные позвонки постепенно увеличиваются в каудальном направлении. Отростки на них слабо развиты, а их концы построены из хрящевой ткани. Такое же строение характерно и для крестцовых позвонков. Они разъединены толстыми прослойками хрящевой ткани. Остистые отростки отделены и прямо поставлены. Между поясничными и крестцовыми позвонками сохраняется значительная подвижность.
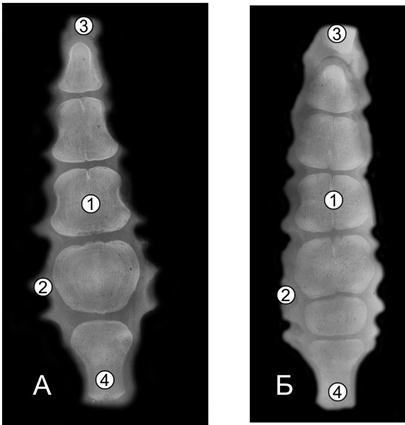
Рис. 107. Рентгенограммы грудины телочек (сутки) пренатальной развитой (А) и пренатальной недоразвитой (Б) групп (Ж. Г. Грабчак, 2004): А – пренатально развитой; Б – пренатально недоразвитой; 1 – губчатая костная ткань; 2 – хрящевая ткань; 3 – рукоятка; 4 –мечевидный отросток.
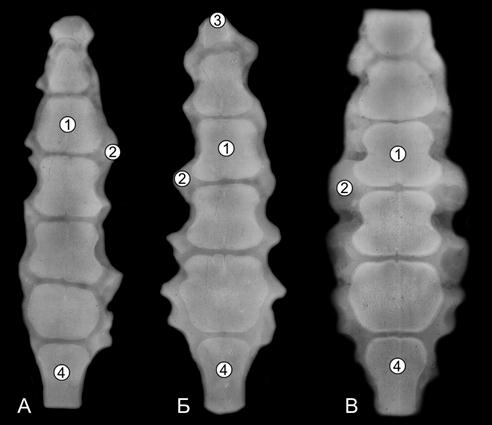
Рис. 108. Рентгенограммы груднини 5- (А), 10-(Б) и 20-(В) телочек (сутки) (Ж. Г. Грабчак, 2004): 1 – губчатая костная ткань; 2 – хрящевая ткань; 3 – рукоятка; 4 – мечевидный отросток.
Скелет хвоста имеет наименьшую абсолютную и относительную массу. Из 18-20 хвостовых позвонков у телят, с высокой жизнеспособностью, только первые 6–7 позвонков имеют три центра окостенения (головки, ямки и диафиза), следующие – лишь два (головки и диафиза), а последние 2-3 позвонка образованы лишь хрящевой тканью [79, 80, 281]. Исследования показывают, что пренатальная особенность остеогенеза хвостовых позвонков может свидетельствовать о росте и развитии не только костной системы, но и всего организма новорожденных (рис. 109-111). Чем больше хвостовых позвонков имеют не только диафизарные (тел), но и эпифизарные (головок и ямок) центры окостенения, тем большую длину имеет хвост, который указывает на интенсивный остеогенез у суточных домашних животных. У недоразвитых животных, кроме того, тела хвостовых позвонков имеют меньшую длину, которая значительно влияет на длину хвоста. С возрастом телят в хвостовых позвонках интенсивно образуются эпифизарные (головки и ямок) центры окостенения.
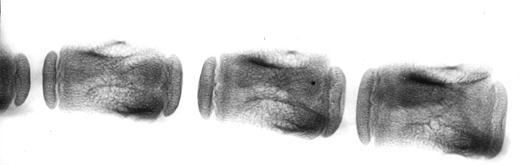
Рис. 109. Рентгенограмма первых трех хвостовых позвонков суточной телочки (П. Н. Гаврилин, 1992): 1 – эпифизарные (головки и ямки); 2 – диафизарные центры окостенения.
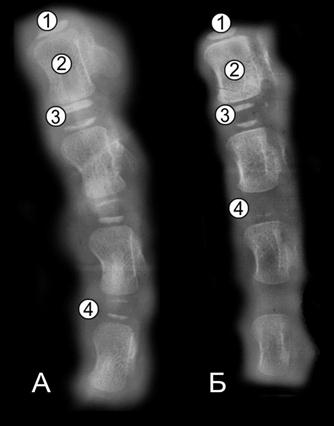
Рис. 110. Рентгенограммы І-ІV хвостовых позвонков телочек (сутки) пренатальной развитой (А) и пренатальной недоразвитой (Б) групп (Ж. Г. Грабчак, 2004): 1 – краниальный центр окостенения; 2 – диафизарный центр окостенения; 3 – каудальный центр окостенения; 4 – хрящевая ткань.
Скелет конечностей новорожденных животных также имеет свои особенности строения. Лопатка содержит значительное количество хрящевой ткани, а в ячейках губчатой костной ткани – остеобластичного костного мозга. Маклаки и седалищные бугры образованы преимущественно гиалиновой хрящевой тканью. Однако, в центре маклака расположено небольшое углубление (ямка), которую используют в качестве один из ориентиров при внутрикостном введении жидких лекарств.
У суточных поросят, с нарушением пренатального остеогенеза лобковые кости почти на 90% образованны из хрящевой ткани. Вследствие этого суставная впадина тазовых костей настолько мелкая, что головка бедренной кости не фиксируется в ней. В таком случае суточные поросята перемещаются в пространстве на грудных конечностях согнутых в запястных суставах, подтягивая тазовые конечности. Такие поросята определяются также сниженной жизнеспособностью [285-287].
Особенностью пренатального роста и развития конечностей у новорожденных животных является более значительное морфофункциональное становление костных органов, по сравнению с мускульными, которые закрепляются на них.
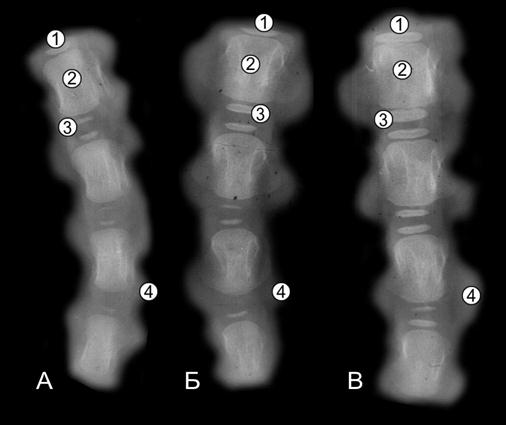
Рис. 111. Рентгенограммы І-ІV хвостовых позвонков 5- (А), 10- (Б) и 20 (В) суточных телочек (Ж. Г. Грабчак, 2004): 1 – краниальный центр окостенения; 2 – диафизарный центр окостенения; 3 – каудальный центр окостенения; 4 – хрящевая ткань.
Костные органы конечностей имеют несколько центров окостенения, развитие которых неодинаково. Наиболее выражена структура компактной и губчатой костной ткани в диафизах трубчатых костей конечностей. Менее выражены центры окостенения в эпифизах (рис. 112). Они не значительные или вообще отсутствуют в апофизах. У новорожденных телят, которые проявляют высокую жизнеспособность, площадь центра окостенения большого вертела бедренной кости колеблется от 3,9-4,7% [76, 81]. У недоразвитых телят и поросят на рентгенограммах костных органов он почти не проявляется (рис. 113).
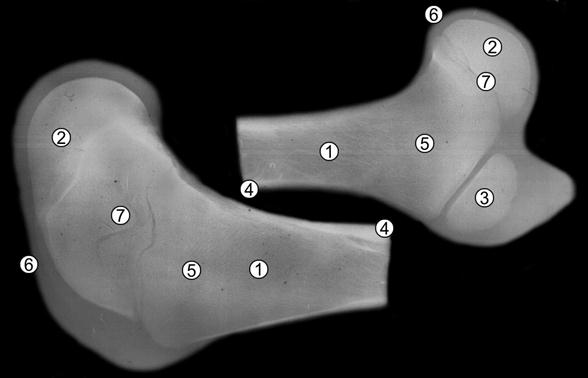
Рис. 112. Рентгенограмма бедренной кости пренатально развитой суточной телочки (Ж. Г. Грабчак, 2004): 1 – костно-мозговой участок; 2 – эпифизарный центр окостенения; 3 – апофизарный центр окостенения; 4 – компактная костная ткань; 5 – губчатая костная ткань; 6 – суставной хрящ; 7 – метафизарный хрящ.
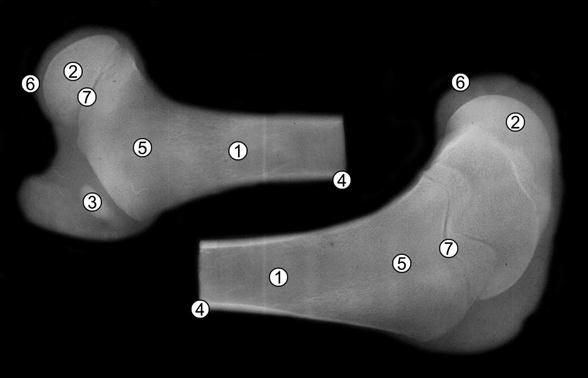
Рис. 113. Рентгенограмма бедренной кости пренатально недоразвитой суточной телочки (Ж. Г. Грабчак, 2004): 1 – костно-мозговая полость; 2 – эпифизарный центр окостенения; 3 – апофизарный центр окостенения; 4 – компактная костная ткань; 5 – губчатая костная ткань; 6 – суставной хрящ; 7 – метафизарный хрящ.
Характерно, что у поросят с живой массой меньше породных показателей, центры окостенения не проявляются даже в головке бедренной кости [234, 287]. При этом у таких животных отмечается нарушение статолокомоторных актов тазовых конечностей.
Кости пясти и плюсны моноэпифизарные – центры окостенения лишь в дистальных эпифизах, которые хорошо развиты. В этих костях красный костный мозг, прежде всего, замещается жировым. Компактная костная ткань приобретает остеонную структуру. Характерно, что в этих костных органах содержится наименьшее количество хрящевой ткани, особенно у жеребят.
Рассечения нативных трубчатых костей грудных и тазовых конечностей у пренатально развитых суточных телочек показывает наличие большого количества губчатой ткани, а в диафизе – толстого слоя компактной (рис. 114, 115). Толщина метафизарных хрящей незначительна. Эпифизы полностью образованы из губчатой костной ткани. Несколько большую толщину имеет суставной хрящ. Однако необходимо отметить, что толщина суставных и метафизарных хрящей уменьшается в дистальном направлении. Они толще в костях стилоподия, по сравнению с метаподиями [155].
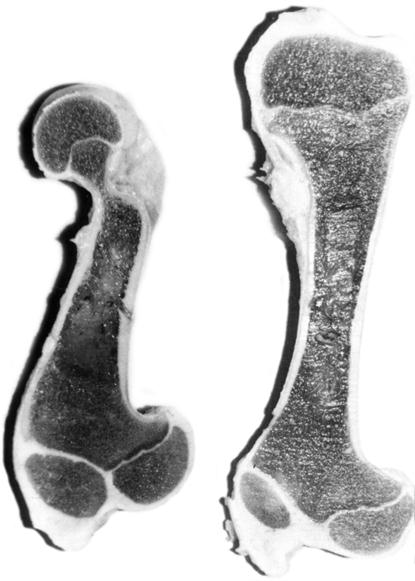
Рис. 114. Рассечение нативных бедренных (1) и плечевой (5) костей неонатального теленка (Б. В. Криштофорова).
У суточных поросят длинные трубчатые кости содержат больше хрящевой ткани, а суставные и метафизарные хрящи имеют относительно большую толщину по сравнению с телятами [234, 287].
У суточных щенков оказываются только диафизарные центры окостенения в длинных трубчатых костях конечностей, тогда как эпифизарные – отсутствуют.
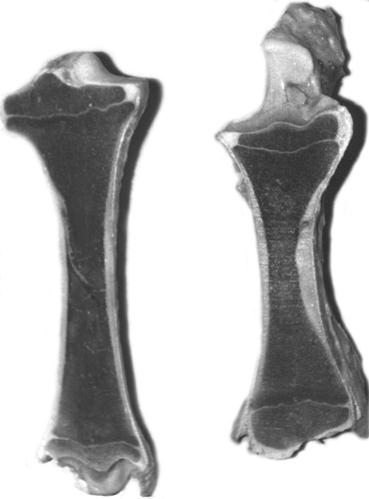

1 2 3 4
Рис. 115. Рассечения нативных большеберцовой (1) и лучевой (2), пястной (3) и плюсневой (4) костей теленка (Б. В. Криштофорова).
Короткие костные органы запястья и заплюсны имеют совсем незначительные центры окостенения, а у щенков они отсутствуют. Площадь окостенения этих костных органов находится в прямой зависимости от интенсивности пренатального остеогенеза и всей костной системы.
Абсолютная масса костных органов грудных конечностей меньше, по сравнению с тазовыми. Абсолютная масса отдельных костных органов суточных телят зависит от породы. Она больше у телят черно-пестрой породы по сравнению с красной степной (табл. 141). Выращивание и содержание коров с использованием дозированного принудительного движения положительно влияет на пренатальный остеогенез, что проявляется увеличением абсолютной массы костных органов (табл. 142). Костные органы проксимальных звеньев имеют большую массу по сравнению с дистальных. В такой же закономерности изменяется относительная масса отдельных костных органов конечностей (табл. 143). Наибольшую относительную массу (к массе костной системы) имеют бедренная и большеберцовая кости, а наименьшую – пястные. Относительная масса груднини достигает всего 1,30%, грудных позвонков – в несколько раз меньше (0,38%), как и 7 ребра (0,30%). У 10-суточных телят относительная масса костных органов незначительно изменяется.
Таблица 141
Абсолютная масса трубчатых костей телочек черно-пестрой степной породы (Б. В. Криштофорова, 1987), г
|
Возраст |
Плечевая |
Бедренная |
Лучевая |
Большеберцовая |
Пястные |
Плюсневые |
|
Новорожденные |
276,62±2,32 |
410,06±4,68 |
227,50±5,01 |
274,50±4,72 |
131,00±3,21 |
153,50± 2,41 |
Таблица 142
Абсолютная масса трубчатых костей телочек красной степной породы (П. Н. Гаврилин, 1992), г
|
Возраст |
Плечевая |
Бедренная |
Лучевая |
Большеберцовая |
Пястные |
Плюсневые |
|
Новорожденные |
237,28± |
356,66± |
212,33± |
263,80± |
127,66± |
140,58± |
|
10 суточные |
276,50-289,66 |
398- |
234,58- |
280,83- |
133,46- |
154,83- |
|
Возраст |
Плечевая |
Бедренная |
Лучевая |
Большеберцовая |
Пястные |
Плюсневые |
|
Новорожденные |
237,28±19,43 |
356,66±24,68 |
212,33±16,34 |
263,80±18,18 |
127,66±7,74 |
140,58±7,53 |
|
10 суточные |
276,50-289,66 |
398-431,50 |
234,58-247,91 |
280,83-310,08 |
133,46-142,05 |
154,83-165,18 |
Таблица 143
Относительная масса и прирост костных органов некоторых телочек красной степной породы (П. Н. Гаврилин, 1992)
|
Кости |
Относительная масса % |
Прирост |
||||
|
Новорожденные |
10-суточные |
Новорожденные |
10-суточные |
Суточный, г |
Относительный % |
|
|
К массе скелета |
К живой массе |
Суточный прирост (г) |
||||
|
Плечевая |
3,20 |
3,14-3,52 |
0,68 |
0,64-0,65 |
3,92-5,32 |
16,50-22,00 |
|
Бедренная |
4,91 |
4,53-5,70 |
1,03 |
0,93-0,97 |
4,23-7,48 |
11,86-20,90 |
|
Лучевая |
2,92 |
2,66-3,01 |
0,61 |
0,54-0,55 |
2,2-3,55 |
10,40-16,70 |
|
Большеберцовая |
3,63 |
3,19-3,77 |
0,76 |
0,65-0,69 |
1,70-4,62 |
6,45-17,54 |
|
Пястные |
1,76 |
1,51-1,72 |
0,37 |
0,30-0,31 |
0,58-1,43 |
4,54-11,27 |
|
Плюсневые |
1,93 |
1,75-2,00 |
0,40 |
0,34-0,36 |
1,42-2,46 |
10,13-17,49 |
|
7 грудной позвонок |
0,38 |
0,42 |
0,079 |
0,077-0,086 |
0,70-0,97 |
25,45-35,27 |
|
7 ребро |
0,30 |
0,36-0,37 |
0,063 |
0,068-0,073 |
0,86-0,96 |
39,52-44,09 |
|
Грудина |
1,30 |
1,43-1,51 |
0,27 |
0,28-0,29 |
3,01-3,11 |
32,00-33,00 |
Объем костных органов конечностей телят красной степной породы также несколько меньше по сравнению с черно-пестрой (табл. 144). Объем костных органов коррелирует с их массой и также снижается в дистальном направлении (бычки черно-пестрой породы: плечевая – 229,00±1,40 см3, бедренная – 343,00±4,63 см3, лучевая – 181,25±4,52 см3, большеберцовая – 220,00±4,61 см3, кости пясти – 94,22±3,87 см3, кости плесни – 110,27±4,85 см3).
Таблица 144
Объем трубчатых костей конечностей телочек красной степной породы, см2 (П. М. Гаврилин, 1992)
|
Возраст |
Плечевая |
Бедренная |
Лучевая |
Большеберцовая |
Пястные |
Плюсневые |
|
Новорожденные |
182,00± |
295,75± |
172,90± |
216,75± |
103,83± |
14,28± |
|
10 суточные |
220,08-233,55 |
310,31-340,15 |
178,55-194,45 |
213,96-238,51 |
97,83-108,20 |
113,25-123,61 |
|
Возраст |
Плечевая |
Бедренная |
Лучевая |
Большеберцовая |
Пястные |
Плюсневые |
|
Новорожденные |
182,00±30,79 |
295,75±4,44 |
172,90±1,20 |
216,75±15,42 |
103,83±6,65 |
14,28±6,47 |
|
10 суточные |
220,08-233,55 |
310,31-340,15 |
178,55-194,45 |
213,96-238,51 |
97,83-108,20 |
113,25-123,61 |
На объем костных органов новорожденных телят влияет розширеность их эпифизов и незначительные выпячивания бугров.
Исследования длины длинных трубчатых костных органов конечностей новорожденных животных показывают, что она больше в тазовых, по сравнению с грудными. Характерно, что у новорожденных телят отношения длины трубчатых костей конечностей к живой массе или массы костной системы имеет определенные закономерности, что можно использовать в качестве тесты при определение пренатального развития животных (табл. 145).
Таблица 145
Отношение длины трубчатых костей к живой массе и абсолютной массе костной массы телочек % (Б. В. Криштофорова, 1987)
|
Название кости |
Отношение к живой массе |
Отношение к скелету |
|
Плечевая |
153,37 |
29,91 |
|
Бедренная |
186,40 |
36,37 |
|
Лучевая |
196,66 |
38,10 |
|
Большеберцовая |
154,91 |
30,21 |
|
Кости пястья |
206,61 |
40,30 |
|
Кости плюсны |
183,99 |
35,69 |
Отношение длины плечевой кости суточных телят несколько меньше (153,37) по сравнению с бедренной (181,40). Однако отношение длины лучевой кости больше (196,66), по сравнению с большеберцовой (154,9). Максимальное отношение к живой массе имеют кости метаподия.
Отношение длины длинных трубчатых костей к абсолютной массе костной системы гораздо меньше при незначительных колебаниях. Бедренная кость суточных телят имеет наиболее стойкие показатели к массе костной системы, которая дает возможность прижизненно определить ее абсолютную массу.
Учитывая живую массу новорожденного теленка и длину трубчатой кости можно установить это отношение и тем самым определить абсолютную массу и особенность пренатального остеогенеза, а также относительную массу всей костной системы. Увеличение этих показателей указывает на недоразвитость костной системы, как и всего организма.
Закономерности изменений соотношений абсолютной массы костных органов к массе скелета и живой массе имеют определенные закономерности. Благодаря этому можно определить абсолютную и относительную массу костной системы, учитывая абсолютную массу бедренной кости (А. Н. Сироткин, 1974):
В = 2,7 + 0,11·Х – 8,78
В – абсолютная масса скелета (кг);
Х – абсолютная масса бедренной кости (г).
Абсолютную массу костной системы можно определить также с учетом живой массы:
В = 2,7 + 0,11·Х
В – абсолютная масса скелета (кг);
Х – живая масса новорожденного животного (кг).
Индекс массивности трубчатых костей конечностей новорожденных телят имеет меньшие показатели, по сравнению с таковыми у взрослых животных. В плечевой кости он достигает 1,58, бедренной – 1,93, лучевой – 1,36, большой голени – 1,30, костях пясти – 0,83; костях плесни – 0,86. У взрослых животных этот показатель увеличивается в два-три разы.
Средняя треть диафиза (как и другие участки) трубчатых костей конечностей имеют определенные периметры, толщину и форму фронтального пересечения. В костных органах стилоподия (плечевая и бедренная), к которым прикрепляется больше всего мышц, фронтальное пересечение имеет почти форму круга.
В костных органах второго звена (зейгоподия) фронтальное сечение в средней трети диафиза изменяется: в лучевой кости он имеет латеро-медиальный овал, тогда как в большеберцовой – дорсо-пальмарный. В костях пясти это также овал в латеро-медиальном направлении, тогда как пясти – равнозначный кругу. На форму фронтального сечения трубчатых костей влияет центр тяжести, которая проходит по грудной или тазовой конечностям.
У новорожденных невзирая на сетчатое строение, компактная костная ткань в каждой трубчатой кости конечностей, имеет определенную толщину (табл. 146).
Следовательно, толщина компактной костной ткани на разных поверхностях трубчатых костей конечностей неодинакова. В плечевой кости она больше на пальмарной; бедренной, лучевой и большеберцовой – на медиальной; а костях пясти и плесни – на латеральной поверхности.
Таблица 146
Толщина компактной костной ткани трубчатых костей конечностей новорожденных бычков черно-пестрой породы, мм (Б. В. Криштофорова, 1987)
|
Поверхность кости |
Плечевая |
Бедренная |
Лучевая |
Большеберцовая |
Кости |
Кости |
|
Дорсальная |
5,25±1,97 |
3,91±1,44 |
2,20±0,86 |
4,01±0,54 |
2,00±0,57 |
3,55±0,87 |
|
Пальмарная (плантарная) |
5,42±0,64 |
3,95±0,83 |
2,44±0,25 |
5,00±0,76 |
2,02±0,42 |
3,75±0,39 |
|
Медиальная |
3,00±0,17 |
6,50±0,24 |
6,85±0,56 |
5,08±0,37 |
3,81±0,44 |
4,80±0,29 |
|
Латеральная |
2,32±0,44 |
5,55±0,85 |
6,35±0,97 |
5,24±0,78 |
4,21±0,38 |
4,59±0,45 |
У новорожденных бычков каждая трубчатая кость конечностей характеризуется показателем развития компактной костной ткани. Этот показатель наибольший в костях зейгоподия (лучевая – 61,80% и большерцовая – 56,82%), несколько меньше метаподия (пясти – 47,69%, плесни – 50,24%) и наименьший – стилоподия (плечевая – 46,69% и бедренная – 47,77%). Следовательно, в костных органах костей стилоподия количество губчатой костной ткани больше по сравнению с костями метаподия.
У новорожденных животных в трубчатых костях конечностей содержится костно-мозговой участок и определено количество губчатой костной ткани, которая расположена в эпифизах и по концам диафиза (табл. 147, 148). В плечевой кости суточных телят губчатая костная ткань занимает 45,61% от всей ее длины, бедренной – 48,05, лучевой – 31,14, большеберцовой – 27, 68%. Наибольшее количество губчатой костной ткани содержится в костях стилоподия.
Ж. Г. Грабчак [76] показывает, что масса костных органов, длина, периметр среднего участка диафиза, высота костномозгового участка диафиза зависит от пренатального развития теленка. Чем больше выражены клинические признаки пренатального недоразвития, тем меньше эти показатели.
У суточных телят определенных размеров занимает костновомозговой участок диафиза трубчатых костей конечностей, который содержит костный мозг с одиночными костными трабекулами (табл. 149). Относительная высота костномозгового участка плечевой кости (к ее длине) у суточных телят достигает 30,70%, в бедренной – 39,27%, в пястной – 47,00%, а плюсневой – 48,68% (наибольшая). С возрастом телят идет незначительный рост костномозгового участка диафиза трубчатых костей конечностей.
Таблица 147
Параметры структурных компонентов бедренной кости телочек красной степной породы (Ж. Г. Грабчак, 2001)
|
Живая |
Масса |
Длина |
Периметр среднего участка диафиза, мм |
Относительная |
Высота |
Ширина |
|
15-30 |
209,90–342,60 |
178–213 |
66–84 |
4,91–16,48 |
36–83 |
8–17 |
|
22-32 |
277,14–386,63 |
198–222 |
72–80 |
2,86–15,92 |
33–80 |
8–19 |
|
26-39 |
300,81–484,52 |
197–229 |
77–86 |
5,43–10,27 |
49–79 |
10–18 |
|
M±m |
337,96±16,96 |
206,95±6,24 |
75,70±1,15 |
9,79±0,.89 |
64,95±3,65 |
15,45±0,75 |
*КМУ – костномозговой участок
Таблица 148
Количество губчатой костной ткани в трубчатых костях конечностей суточных телят (% от длины кости) (Б. В. Криштофорова, 1987)
|
Участок кости |
Плечевая |
Бедренная |
Лучевая |
Большеберцовая |
Кости |
Кости |
|
Проксимальный эпифиз |
10,05 |
7,51 |
7,70 |
9,29 |
– |
– |
|
Дистальный эпифиз |
7,69 |
9,99 |
8,11 |
4,81 |
15,25 |
13,80 |
|
Проксимальна участок диафиза |
17,24 |
17,71 |
9,16 |
10,01 |
10,95 |
14,35 |
|
Дистальный участок диафиза |
19,98 |
15,83 |
19,53 |
10,00 |
18,64 |
19,86 |
|
Всего в % |
45,61 |
48,05 |
31,14 |
27,68 |
19,16 |
20,38 |
Таблица 149
Высота костно-мозгового участка диафиза трубчатых костей конечностей телочек красной степной породы % (П. Н. Гаврилин, 1992)
|
Название |
Новорожденные |
10-суточные |
|
Плечевая |
53,00±2,10 |
56,11-56,47 |
|
30,70 |
30,78-30,96 |
|
|
Бедренная |
83,13±4,60 |
81,91-87,99 |
|
39,27 |
39,80-39,89 |
|
|
Лучевая |
69,50±2,30 |
69,15-81,87 |
|
40,60 |
40,02-40,42 |
|
|
Большеберцовая |
91,07±5,60 |
93,00-93,38 |
|
42,76 |
42,35-42,78 |
|
|
Пястная |
75,30±0,20 |
78,25-78,60 |
|
47,00 |
47,39-47,63 |
|
|
Плюсневая |
86,94±4,40 |
89,24-92,10 |
|
48,68 |
48,46-48,69 |
Определение плотности костных органов (их отдельных участков) является одним из показателей, который позволяет провести сравнение между собой невзирая на разнообразие их анатомического строения. Необходимо отметить, что плотность костного мозга, который содержится в диафизе трубчатых костей конечностей суточных телят очень низкая и колеблется в пределах 0,93-0,97 г/см3 (более малая единице, табл. 150). У взрослых животных плотность костного мозга еще меньше (0,84-0,86 г/см), что предопределенно наличием большого количества жировых клеток.
У новорожденных животных плотность целых костных органов наименьшая. Однако плотность костных органов осевого скелета еще более малая (табл. 151). Плотность позвонков несколько большая (1,092±0,007 г/см3) по сравнению с грудиной (1,086±0,02 г/см3) и, особенно, ребрами (0,043±0,005г/см3).
Плотность трубчатых костей конечностей значительно больше, по сравнению с позвонками (табл. 152). Плотность костных органов пальцев у суточных телят изменяется с обратной закономерностью – она растет в проксимальном направлении (табл. 153). Плотность копытцевой кости меньше по сравнению с путовой.
Плотность разных участков костных органов суточных животных, особенно трубчатых костей конечностей, также неодинакова, что определяется соотношением губчатой и компактной костной ткани (табл. 154). Плотность костей стилоподия меньше, чем метаподия. При этом наибольшая плотность компактной костной ткани отмечается в среднем участке диафиза трубчатых костей, а наименьшая – в эпифизе, где количество хрящевой ткани наибольшее. Промежуточное значение плотности имеют проксимальные и дистальные участки эпифиза, в которых постепенно нарастает толщина компактной костной ткани и уменьшается количество хрящевой.
Таблица 150
Плотность костного мозга костно-мозгового участка диафиза трубчатых костей бычков черно-пестрой породы, г/см3 (Б. В. Криштофорова, 1987)
|
Возраст |
Плечевая |
Бедренная |
Лучевая |
Большеберцовая |
Пястные |
Плюсневые |
|
Новорожденные |
0,99±0,033 |
0,989±0,038 |
0,986±0,063 |
0,988±0,051 |
0,979±0,46 |
0,976±0,027 |
|
30 суточные |
0,95–0,973 |
0,948–0,965 |
0,941–0,958 |
0,939–0,953 |
0,933–0,949 |
0,938–0,951 |
Таблица 151
Масса, объем и плотность костей грудного сегмента телочек красной степной породы (П. Н. Гаврилин, 1992)
|
Возраст |
Кость |
Масса, г |
Объем, см3 |
Плотность, г/см3 |
|
Новорожденные |
7 грудной позвонок |
27,50±4,60 |
25,17±0,02 |
1,092±0,007 |
|
7 ребро |
21,86±2,83 |
20,93±2,62 |
1,043±0,005 |
|
|
Грудина |
94,34±5,90 |
86,66±5,90 |
1,086±0,002 |
|
|
10 суточные |
7 грудной позвонок |
34,50–37,30 |
27,73–29,45 |
1,246–1,263 |
|
7 ребро |
30,50–31,50 |
27,29–28,04 |
1,150–1,154 |
|
|
Грудина |
124,53–125,50 |
106,79–107,37 |
1,164–1,167 |
Таблица 152
Плотность трубчатых костей конечностей телочек красной степной породы, г/см3 (П. М. Гаврилин, 1992)
|
Возраст |
Плечевая |
Бедренная |
Лучевая |
Большеберцовая |
Пястная |
Плюсневая |
|
Новорожденные |
1,194±0,002 |
1,205±0,04 |
1,226±0,003 |
1,212±0,001 |
1,241±0,006 |
1,248±0,006 |
|
10 суточные |
1,237–1,256 |
1,273–1,215 |
1,274–1,313 |
1,299–1,312 |
1,319–1,363 |
1,336–1,366 |
Таблица 153
Плотность костей пальцев телочек красной степной породы, г/см3 (В. В. Яшина, 1994)
|
Возраст |
Конечность |
Пальцы |
Путовая |
Венечная |
Копытцевая |
|
Новорожденные |
Г |
3 |
1,197±0,01 |
1,168±0,01 |
1,189±0,02 |
|
4 |
1,181±0,01 |
1,160±0,02 |
1,143±0,02 |
||
|
Т |
3 |
1,90±0,01 |
1,167±0,02 |
1,166±0,01 |
|
|
4 |
1,90±0,01 |
1,172±0,01 |
1,139±0,02 |
||
|
10 суточные |
Г |
3 |
1,195–1,217 |
1,194–1,207 |
1,151–1,160 |
|
4 |
1,202–1,227 |
1,182–1,214 |
1,189–1,251 |
||
|
Т |
3 |
1,213–1,219 |
1,197–1,202 |
1,59–1,166 |
|
|
4 |
1,205–1,226 |
1,200–1,230 |
1,180–1,104 |
Таблица 154.
Плотность разных участков трубчатых костей конечностей новорожденных бычков черно-пестрой породы, г/ см3 (Б. В. Криштофорова, 1987)
|
Участки костей |
Название кости |
|||||
|
Плечевая |
Бедренная |
Лучевая |
Большеберцовая |
Кости пясти |
Кости |
|
|
Проксимальный эпифиз |
1,060±0,21 |
1,109±0,19 |
1,077±0,46 |
1,086±0,02 |
– |
– |
|
Дистальный Эпифиз |
1,136±0,06 |
1,006±0,05 |
1,086±0,03 |
1,084±0,06 |
1,094±0,03 |
1,091±0,16 |
|
Проксимальна участок диафиза |
1,143±0,02 |
1,149±0,03 |
1,164±0,03 |
1,157±0,04 |
1,123±0,05 |
1,196±0,04 |
|
Дистальный участок диафиза |
1,123±0,03 |
1,115±0,04 |
1,172±0,03 |
1,152±0,04 |
1,172±0,03 |
1,152±0,04 |
|
Средний участок диафиза |
1,253±0,004 |
1,335±0,01 |
1,321±0,01 |
1,377±0,002 |
1,417±0,003 |
1,439±0,001 |
|
Всей трубчатой кости |
1,20±0,04 |
1,19±0,02 |
1,25±0,001 |
1,24±0,001 |
1,28±0,024 |
1,29±0,02 |
Наибольшая плотность минеральных веществ оказывается в компактной костной ткани, а наименьшая – в губчатой.
Плотность костных органов, их отдельных участков положительно коррелирует с минерализацией: чем меньшая плотность, тем меньше содержится минеральных веществ. Наибольшее количество минеральных веществ содержится в компактной костной ткани диафиза костей плюсны (табл. 155), а наименьшая – в плечевой. В эпифизах трубчатых костей конечностей суточных телят содержится наименьшее количество минеральных веществ (6,73±0,06%). Их количество несколько растет в проксимальном и дистальном участках диафиза.
Таблица 155
Содержание минеральных веществ в разных участках трубчатых костных органов конечностей бычков черно-пестрой породы % (Б. В. Криштофорова, 1987)
|
Участки костных органов |
Название кости |
|||||
|
Плечевая |
Бедренная |
Лучевая |
Большеберцовая |
Пясти |
Плюсны |
|
|
Проксимальный |
6,73±0,06 |
7,56±0,93 |
9,93±1,29 |
9,78±1,10 |
– |
– |
|
Дистальный эпифиз |
7,43±1,11 |
5,69±0,89 |
8,63±0,89 |
7,21±0,94 |
9,63±0,83 |
9,63±0,67 |
|
Проксимальна участок диафизу |
9,27±1,23 |
9,33±1,41 |
10,46±1,17 |
10,87±1,33 |
11,29±1,09 |
9,81±0,94 |
|
Дистальный участок диафиза |
9,33±0,98 |
8,86±1,03 |
9,56±1,23 |
9,69±1,15 |
10,49±1,03 |
11,17±1,21 |
|
Средний участок диафиза |
17,36±1,05 |
17,89±1,23 |
18,69±0,94 |
18,46±0,83 |
18,84±1,15 |
18,93±1,21 |
Незначительная минерализация костных органов у новорожденных телят нуждается в интенсивном поступлении с кормом минеральных веществ. По мнению В. В. Митюшина [219] потребность в минеральных веществах организмом новорожденных обеспечивается макро - и микроэлементами молозива, которое определяется высоким процентом их потребления (табл. 156).
Таблица 156
Потребление минеральных веществ организмом телят в период новорожденности (В. В. Митюшин, 1979)
|
Название |
Усвоено в % от принятого |
||
|
1 сутки |
2-6 суток |
6-10 суток |
|
|
Вода |
10,0 |
0,3 |
– |
|
Натрий |
24,2 |
40,4 |
39,3 |
|
Калий |
69,3 |
23,9 |
17,9 |
|
Кальций |
98,0 |
82,2 |
94,0 |
|
Магний |
76,9 |
62,9 |
61,6 |
|
Хлор |
45,7 |
15,9 |
5,2 |
|
Фосфор |
92,4 |
45,4 |
36,4 |
У телят в период новорожденности, наибольшая потребность организма отмечается в кальции, потребление которого достигает 94%, а фосфору – 36,4%. Данный факт является доказательством, что в этот период не только проходит рост и развитие костной системы, но и ее трансформация, которая связана с усилением функций гемопоэза и иммуногенезу. Доказано, что выгульное содержание телят усиливает потребление минеральных веществ. В костной ткани депонируется кальция на 1,33-1,49%, фосфору – 0,69-0,78% больше по сравнению с таковыми показателями телят, которые выращивают в клетках. Установлено, что соответственно росту и развитию телят, в первые сутки их жизни, необходимо 27 г кальцию и 18 г фосфору. Такая же потребность в макро - и микроэлементах отмечается у жеребят, поросят и ягнят.
Костные органы образованы из определенных тканевых компонентов, соотношение которых влияет на функциональную их активность. Взаимоотношение тканевых компонентов в костных органах разное, но оно, в первую очередь, зависит от пренатального развития новорожденных млекопитающих.
От соотношения костной и хрящевой ткани в костных органах зависит количество остеобластического и красного костного мозга. Доказана зависимость соотношения количества хрящевой и костной ткани от живой массы новорожденного теленка (табл. 157). Автор [100] досказывает, что чем меньшая живая масса новорожденного теленка, тем более в костной системе содержится хрящевой ткани (15%). С ростом живой массы суточных телят увеличивается количество костной ткани (87,44%) и уменьшается хрящевой (12,56%).
Таблица 157.
Масса костной и хрящевой ткани в скелете новорожденных телят (Н. А. Зикин, 1980)
|
Живая масса, г |
Костная ткань |
Хрящевая ткань |
||
|
Г |
% |
Г |
% |
|
|
27100 |
7364,89 |
65,00 |
1299,71 |
15,00 |
|
28300 |
7375,23 |
84,91 |
1311,55 |
15,09 |
|
35600 |
7795,16 |
87,07 |
1150,61 |
12,93 |
|
36700 |
7845,63 |
87,44 |
1127,52 |
12,56 |
По данным П. Н. Гаврилина [65, 66, 69] в отдельных костных органах новорожденных телят количество костной, хрящевой ткани и костного мозга также зависит от биофизических нагрузок. В костных органах конечностей преобладает костная ткань и красный костный мозг, тогда как в осевом хрящевая ткань и остеобластический костный мозг (табл. 158). Так костной ткани в позвоночном конце первого ребра содержится 28,05±0,97%, в 7-33,54±0,18% и в 13-28,89±0,53%. В разных позвонках количество костной ткани, как и хрящевой неодинаково. В первом грудном позвонке ее количество достигает 31,50±0,49%, в поясничном – 31,90±0,69% и 2 хвостовом – 23,07±0,69%, а 10- хвостового всего 22,58±0,60%. С уменьшением количества костной ткани растет количество хрящевой.
Таблица 158.
Относительное количество структурных компонентов некоторых костных органов осевого скелета новорожденных телочек красной степной породы % (П. Н. Гаврилин, 1998)
|
Название |
Хрящевая ткань |
Костная ткань |
Костный мозг |
||
|
Грудина (3 сегмент) |
26,00±0,96 |
21,28±0,36 |
49,60±0,50 |
||
|
Ребро (головка, |
13 |
33,81±0,46 |
28,89±0,53 |
32,83±0,50 |
|
|
7 |
15,31±0,44 |
33,54±0,18 |
46,35±0,57 |
||
|
1 |
26,72±0,61 |
28,05±0,97 |
41,78±0,22 |
||
|
Позвонок (тело) |
Хвостовой |
13 |
37,05±0,14 |
19,05±0,48 |
38,98±0,40 |
|
10 |
30,94±0,82 |
22,58±0,43 |
41,46±0,49 |
||
|
7 |
28,21±0,76 |
22,51±0,60 |
43,52±0,67 |
||
|
2 |
24,01±0,46 |
23,07±1,00 |
48,57±0,89 |
||
|
Поясничный |
6 |
22,14±1,00 |
31,90±0,69 |
42,35±0,46 |
|
|
Грудной |
13 |
22,37±1,06 |
30,29±0,47 |
42,70±0,40 |
|
|
7 |
20,20±1,15 |
31,38±0,70 |
49,03±0,52 |
||
|
1 |
19,45±1,39 |
31,50±1,05 |
42,80±0,32 |
||
|
Шейный |
3 |
20,24±0,57 |
31,25±0,49 |
42,04±0,36 |
|
|
Клиновидная (тело) |
7,19±0,13 |
29,66±0,44 |
50,67±0,49 |
Большое количество хрящевой ткани в костных органах указывает на пренатальную недоразвитость животных, которая являются одной из причин их заболеваемости (до 100%) с нередким летальным концом (до 50%) [155-160].
Количество хрящевой ткани в костных органах предопределяется наличием гиалинового хряща. В трубчатых костях новорожденных животных он образует суставные и метафизарные хрящи, которые имеют неодинаковую толщину (табл. 159). В плечевой кости толщина проксимального суставного хряща больше, по сравнению с дистальным, а в бедренной, напротив, более малая. У суточных телят толщина проксимального суставного хряща большеберцовой кости максимальна. В костях пясти и плесни толщина проксимального суставного хряща незначительно отличается от дистального. Колебания толщины метафизарных хрящей трубчатых костей значительно меньше чем суставных. Однако метафизарный хрящ толще в костях стилоподия. по сравнению с таковым метаподия [155].
Таблица 159
Толщина суставных и метафизарных хрящей трубчатых костных органов конечностей бычков черно-пестрой породы, мм (Б. В. Криштофорова, 1987)
|
Участки костей |
Название кости |
|||||
|
Плечевая |
Бедренная |
Лучевая |
Большеберцовая |
Кости пясти |
Кости |
|
|
Проксимальный |
2,44±0,43 |
2,26±0,26 |
1,97±0,17 |
3,08±0,03 |
1,80±0,56 |
2,31±0,64 |
|
Дистальный |
2,21±0,13 |
5,55±1,03 |
2,06±0,83 |
1,04±0,44 |
2,16±0,51 |
2,15±0,30 |
|
Проксимальный |
1,31±0,76 |
1,52±0,43 |
1,33±0,50 |
1,45±0,23 |
– |
– |
|
Дистальный |
1,08±0,33 |
1,25±0,21 |
1,01±0,24 |
0,98±0,17 |
1,10±0,84 |
1,35±0,47 |
Исследования показывают, что относительное количество костной и хрящевой ткани в трубчатых костях (их участках) конечностей суточных телочек неодинаково (табл. 160). Однако, закономерность позитивной взаимосвязи костной ткани и костного мозга присуща для каждого костного органа. В проксимальном эпифизе плечевой кости содержится 31,40±0,37% костной ткани, костного мозга – 50,72±0,26%, тогда как в дистальном – 34,67±0,15% и 52,03±0,28%. В эпифизах бедренной кости количество костной ткани и костного мозга меньше. В дистальном эпифизе пястных костей у суточных телят содержится наименьшее количество хрящевой ткани, как костного мозга [134].
Таблица 160
Относительное количество структурных компонентов в эпифизах трубчатых костей новорожденных телочек красной степной породы % (П. Н. Гаврилин, 1998)
|
Кость |
Эпифизы |
Хрящевая ткань |
Костная ткань |
Костный мозг |
|
Плечевая |
Прокс. |
16,60±0,55 |
31,40±0,37 |
50,72±0,26 |
|
Дист. |
10,90±0,28 |
34,67±0,15 |
52,03±0,28 |
|
|
Лучевая |
Прокс. |
16,83±0,60 |
31,69±0,54 |
49,26±0,29 |
|
Дист. |
14,55±0,18 |
34,86±0,16 |
49,69±0,25 |
|
|
Пястные |
Дист. |
6,25±0,36 |
38,22±0,75 |
52,62±0,37 |
|
Бедренная |
Прокс. |
19,67±0,91 |
30,93±1,60 |
48,05±0,42 |
|
Дист. |
30,30±0,28 |
31,04±0,47 |
46,94±0,20 |
|
|
Большеберцовая |
Прокс. |
16,50±0,53 |
24,32±0,07 |
57,16±0,44 |
|
Дист. |
7,90±0,31 |
37,42±0,32 |
52,77±0,24 |
|
|
Плюсневая |
Дист |
7,02±0,28 |
38,69±0,34 |
53,18±0,30 |
Следовательно, наибольшая толщина хрящей отмечается в проксимальном эпифизе костей стилоподия, за исключением суставного хряща дистального эпифиза бедренной кости. Метафизарный хрящ самый толстый в костях стилоподия и самый тонкий – большеберцовый [134].
Определена закономерность толщины хрящей, соотношения тканевых компонентов характерная и для костей пальцев грудных и тазовых конечностей (табл. 161, 162). В совокупности толщина суставных и хрящей путовой и венечных костей кое-что преобладает толщину метафизарных. Кроме этого, остатки хрящевой ткани оказываются в трабекулах губчатой костной ткани примитивного строения (особенно в зонах роста). В отличие от других костных органов, толщина суставных и метафизарных хрящей путовой и венечных костей пальцев у телят 10 суточного возраста значительно уменьшается [357, 358].
Таблица 161
Толщина хрящей путовой кости телочек красной степной породы, мкм (В. В. Яшина, 1994)
|
Возраст |
Конечность |
Пальцы |
Суставной |
Метафи-зарный проксималь- Ный |
Суставной |
|
Новорожденные |
Г* |
3 |
780±0,05 |
480±0,03 |
850±0,05 |
|
4 |
830±0,05 |
450±0,03 |
880±0,09 |
||
|
Т |
3 |
700±0,07 |
500±0,02 |
880±0,09 |
|
|
4 |
780±0,08 |
530±0,03 |
1050±0,05 |
||
|
10 суточные |
Г |
3 |
883–867 |
417–483 |
767±1017 |
|
4 |
700–867 |
383–467 |
600±767 |
||
|
Т |
3 |
750–883 |
383–417 |
717±1017 |
|
|
4 |
700–750 |
383–433 |
700±783 |
*Г и Т – грудные и тазовые конечности
Таблица 162.
Толщина хрящей венечной кости телочек красной степной породы, мкм (В. В. Яшина, 1994)
|
Век, |
Конечность |
Пальцы |
Суставной |
Метафизарный проксимальный |
Суставной |
|
1 |
Г* |
3 |
940±0,08 |
460±0,04 |
1000±0,13 |
|
4 |
980±0,05 |
460±0,01 |
900±0,12 |
||
|
Т* |
3 |
950±0,11 |
450±0,00 |
900±0,07 |
|
|
4 |
1000±0,07 |
500±0,04 |
900±0,07 |
||
|
10 |
Г |
3 |
767–983 |
283±400 |
717–967 |
|
4 |
667–750 |
383–467 |
700–917 |
||
|
Т |
3 |
767–983 |
417–433 |
667–783 |
|
|
4 |
733–867 |
367–383 |
683–733 |
* Г – грудная, Т – тазовая
Наибольшая разница строения костных органов новорожденных животных регистрируется на микроскопическом уровне. Во всех костных органах новорожденных продуктивных животных компактная и губчатая костная ткань ретикулофиброзная (незрелая). Компактная костная ткань образована костными пластинками, которые соединяются между собой, образовывая сетку. Остеоцити имеют звездчатую форму и значительно увеличены в размерах. По направлению к эпифизам компактная костная ткань истончается, количество трабекул уменьшается и у них появляются включения из гиалинового хряща. В пространствах между трабекулами содержатся остеобласты и соединительная ткань, которая содержит кровеносные сосуды.
Губчатая костная ткань примитивного строения. Трабекули, которые ее образуют разветвляются незначительно. Характерно, что почти во всех трабекулах содержится то или иное количество хрящевой ткани. В эпифизах губчатая костная ткань образуется в зонах роста суставных хрящей, тогда как в диафизе – метафизарных. В проксимальных и дистальных участках диафиза костных органов у новорожденных продуктивных животных рядом с образованием губчатой костной ткани, отмечается ее разрушение, которое предопределяет увеличение высоты участка заполненного костным мозгом. Разрушение трабекул губчатой костной ткани происходит без видимой ее фрагментации, а с характерным лизисом и последующей трансформацией в остеобластический костный мозг. Можно допустить, что такое явление вызывается недостаточным поступлением с кормом питательных веществ необходимых для становления костными органами функции универсального гемопоэза при замещении утробных форменных элементов крови на дефинитивные, характерные для новорожденного периода.
Во всех костных органах рядом с определенным количеством остеобластичного костного мозга содержится красный (кроветворный), что представлено в разделе особенностей органов кроветворения и иммунной защиты. В костных органах осевого скелета, за исключением хвостовых позвонков, оказываются лишь одиночные жировые клетки, тогда как в костях конечностей и, особенно их среднему участку диафиза, содержится желтый костный мозг с вкраплением островков гемопоэтического (см. раздел 3.1).
Разница в строении зон роста отмечается в трубчатых костях новорожденных животных. Зону роста костных органов образуют три основных компонента: гиалиновый хрящи, СХКТ, в которых расположено микроциркуляторное русло. Гиалиновая хрящевая ткань образует суставные и метафизарные хрящи. Суставные хрящи у всех новорожденных животных как и у взрослых животных не имеют надхрящницы имеют достаточно толстую основную зону (зону покоя с незначительным выражением размножения хондроцитов), в которой содержатся одиночные хондробласти и однородное межклеточное вещество. Незначительно меньшего развития достигает зона хрящевых колонок и зона кальцификации суставного хряща. В последней проходит интенсивное разрушение хрящевого матрикса в результате чего образуюсь расширенные лакуны, заполненные кровеносными капиллярами и бесструктурным веществом оксифильного характера. На поверхности остатков хрящевых колонок образуется остеоид, который размещается тонкими пластинками.
СХКТ суставного хряща образована тонкими, небольшой длины трабекулами ретикулофиброзной костной ткани, которая содержится на остатках хрящевых трабекул. Овальной формы ячейки заполнены гемокапилярами и осеобластическим костным мозгом, в котором регистрируются одиночные скопления кроветворных клеток. Между капиллярами оказывается не значительное количество оксифильной бесструктурного вещества. СХКТ суставного хряща без четких границ переходит в первичную губчатую костную ткань, в которой отмечается увеличение костной ретикуло-фиброзной ткани, расширения костно-мозговых ячеек и уменьшения количества хрящевой ткани. Трабекули губчатой костной ткани имеют преимущественно радиальное направление. Только на границе из метафизарным хрящом они направлены косо горизонтально, образовывая эпиметафизарную СХКТ.
Метафизарный хрящ в костных органах имеет однотипное строение, для которого характерно отсутствие морфологической полярности. Со стороны эпифизов на границе с СХКТ происходят интенсивные процессы образования костной ткани. Это приводит к раслоению хряща в виде небольших образований лакун на границе которых содержится один или два слоя хондроцитов в состоянии деструкции. Между ними содержится межклеточное вещество в которой оказываются волокнистые структуры. В несколько более глубоких прослойках хондробласти уменьшаются в размерах и увеличивается площадь межклеточного вещества. По направлению к диафизу в метафизарном хряще костных органов новорожденных животных оказываются все зоны. Особенно хорошо развитая основная и зона хрящевых колонок. В последней содержится 6-8 рядов хондроцитов. В зоне кальцификации наблюдается разрушение хряща и образование тонких наслоений остеоида. Вместе с диафизарной СХКТ образуется ростковая зона, которая обеспечивает рост диафиза. Щели между трабекулами содержат продольные капилляры, заполненные эритроцитами. На поверхности костных трабекул в виде монослоя расположены остеобласты.
Диаметафизарная СХКТ имеет значительно большую толщины, по сравнению с суставным хрящом. Она образована тонкими, заостренными на концах полосками фиброзно-ретикулярной ткани на трабекулах первичной губчатой костной ткани, содержащих значительное количество хрящевой ткани и находятся в состоянии разрушения. Ячейки заполнены удлиненными капиллярами и остеобластами. Диаметафизарная СХКТ обеспечивает интенсивный рост диафиза трубчатых костей в длину, которая отражается на длине конечностей продуктивных животных [134-161].
Необходимо отметить, что у новорожденных продуктивных животных центры синостоза, пронизывают всю толщу хряща от эпифиза к диафиза. Эти одиночные, очень тонкие иглообразные синостози встречаются не только в метафизарных хрящах костей конечностей, но и в позвонках. Они отсутствуют лишь в грудине. Синостози образованы тонкими пластинками остеоида, который окружает кровеносный сосуд. Наличие синостозов, которые пронизывают метафизарный хрящ отмечали П. М. Гаврилин [65, 69], Ж. Г. Грабчак [77,78], Б. В. Криштофорова [134] и В. В. Яшина [357]. Синостоз костных органов неонатальных животных имеет, возможно, временный характер и предопределяется нарушением остеогенеза в пренатальный период онтогенеза. Однако, на наш взгляд это явление нуждается в проведении последующих исследований. Необходимо лишь отметить, что этот фактор в сочетании с содержанием животных в неблагоприятных условиях является одним из факторов нарушения структуры и функции костной системы у взрослых животных.
Особенности структуры и функции костных органов зависят от состояния и архитектоники кровеносных сосудов, которые определяют структуру костной ткани и костного мозга. В зонах роста, где преобладает Ретикуло-фиброзная костная ткань и остеобластический костный мозг [77], кровеносные капилляры имеют значительную длину, заполненные эритроцитами, а их стенка образована эндотелиоцитами. Капилляры слепо заканчиваются в пограничной зоне кальцифицированного хряща. Такие капилляры контактируют с остеобластами. С появлением скоплений кроветворных клеток капилляры образуют полигональные сети. Среди них оказываются редко расположенные артерии мышечного типа.
В целом интраосальные кровеносные сосуды имеют характерную архитектонику и строение, которое предопределено биомеханической и кроветворной функциями. В костных органах, где преобладает губчатая костная ткань вены образуют параартериальные сплетение или же проходят самостоятельно и выходят на поверхность в участке закрепления суставной капсулы. В проксимальном и дистальном участках трубчатых костных органов, вены перед выходом за границу компактной костной ткани разветвляются на более мелких (иногда даже ближе к венулам), а затем опять соединяются и образуют экстарорганное венозное сплетение. Питательная диафизарная артерия и вены имеют наименьший поперечник в костях метаподия и наибольший – зейгоподия. При этом, питательная вена в 2-3 раза тоньше артерии. В плечевой кости она может отсутствовать в 85% случаев. Толщина диафизарных артерий и вен увеличивается в проксимальном направлении размещения длинных трубчатых костей конечностей. Диафизарные вены и артерии образуют целую систему древовидно разветвленных сосудов. Конечные ветвления артерий и начало венозной сетки имеют место в ростковой зоне, а стволы их проходят в средней трети диафиза. От венозного ствола ответвляется питательная вена, которая вливается в вены окружающих мягких тканях. Основное течение крови проходит через эпифизарные артерии и вены [78, 133].
Детальные исследования показывают, что диафизарная костно-мозговая вена образует нисходящие и восходящие ветви. При этом в костях метаподия (пясти и плюсне) они практически одинаковые, зейгоподия (лучевая и бодьшеберцоваяголень) значительно преобладает нисходящая, а стилоподия (плечевая и бедренная) – восходящая.
Взаимоотношения диафизарно-костно-мозговых вен имеют характерные особенности. Прежде всего, диафизарная костно-мозговая артерия, сопровождая вену, образует паравенозное артериальное сплетение развитое больше в тех костных органах, где меньшая микротвердость, а упругие деформации больше [133, 333, 334].
Можно допустить, что спиралеобразные размещение ветвей, как самой диафизарной артерии, выполняет функцию своеобразного жома и этим самым способствует течения крови из интраосальных вен, компенсируя недостаточные упругие деформации в данном участке костного органа (рис. 116). Характерно, что архитектоника взаимоотношений диафизарной артерии и вены почти во всех трубчатых костных органах одинаковая. Однако, у суточных телят более развитая в плечевой кости по сравнению с бедренной (рис. 117, 118). Двигательная активность животных в первые 20 суток жизни также побуждает развитие диафизарных артерий, вен и их взаимоотношений (рис. 119).
В костных органах, особенно в губчатой костной ткани, интраосальные вены образуют сплетения, которое предопределяют течение крови в разных направлениях при условии изменения действия сил упругих деформаций (рис. 120).
Рис. 116. Фронтальный срез диафизарной артерии и вены паравенозного артериального сплетения плечевой кости теленка. Гематоксилин и эозин, МБИ-6, 10х8 (Б. В. Криштофорова, 1974).
Рис. 117. Паравенозные артериальные сплетения в диафизе плечевой кости теленка. Интраосальная инъекция вен тушью на 3% растворе желатина. МБС-10, 8х2 (Б. В. Криштофорова, 1974).
.
Рис. 118. Паравенозне артериальное сплетение в диафизе бедренной кости теленка (сутки). Интраосальная инъекция вен тушью на 3% растворе желатина. МБС-10, 8х2 (Б. В. Криштофорова, 1974).
Рис. 119. Паравенозне артериальное сплетение в диафизе бедренной кости теленка (20 суток). Интраосальная инъекция вен тушью на 3% растворе желатина. МБС-10 8х10 (Б. В. Криштофорова, 1974).
.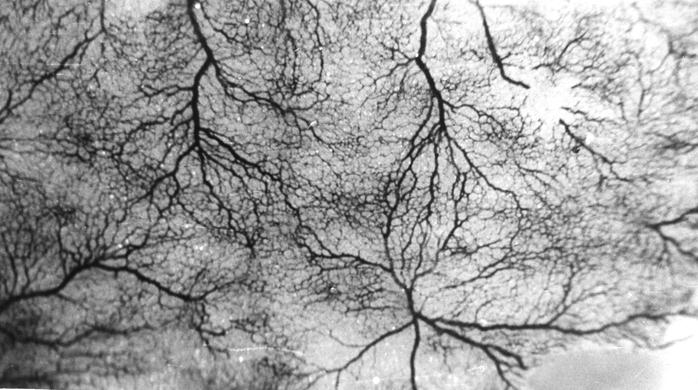
Рис. 120. Вены и сосуды микроциркуляторного русла диафиза бедренной кости теленка (сутки). Интраосальная инъекция тушью на 3% растворе желатина. МБС-10, 8х2 (Б. В. Криштофорова, 1974).
Интраосальные вены образуются слиянием вен и следуют к диафизарной вене (рис. 121, 122). Стенка диафизарной вены очень тонкая. При импрегнации свинцом сосудов диафиза просматривается эндотелий, который содержит округлой формы ядро (рис. 123).
Заслуживает внимания структура посткапиляров и венул. Они также имеют тонкую стенку и отличаются лишь увеличением поперечника (рис. 124). Эндотелиоциты содержат достаточно большое округлое ядро (рис. 125).
В губчатой костной ткани, где больше всего выражены упругие деформации, вены, напротив, оплетают артерии, образовывая параартериальное венозное сплетение. Кроме того, толщина стенки артерий больше ее просвета, тогда как вены тонкостенные и преимущественно округлой формы.
Расположение артериальных и венозных магистралей в губчатой кости эипфизов– горизонтальное, а в диафизе – параллельно длине костного органа.
Тип разветвления артерий и вен в каждом костном органе, в определенном его участке имеет свои особенности. Основной тип разветвления древовидный, однако встречается кустообразный и даже паукообразный. В губчатой костной ткани он больше рассыпной, а вены образуют полигональные сетки, что способствуют течению крови в разных направлениях.
В костных органах первой и второй фаланги (путовая и венечная) конечной артерии и вены образуют разветвление ветвей, как и в длинных трубчатых костях. В третьей фаланге, вокруг внутриканальных артерий, тонкостенные вены образуют характерные пещеристые тела. Кроме этого, тонкостенные вены окружены эластичным каркасом и коллагеновыми волокнами, которые проникают в окружающую ткань. Исследования показывают, что интраосальные сосуды копытцевых костей принимают участие в гемодинамике всего пальца [192, 193]. Взаимоотношение суммарного поперечника артерий и вен в костных органах имеет свои характерные особенности, как и их относительная площадь и количество (табл. 163, 164).
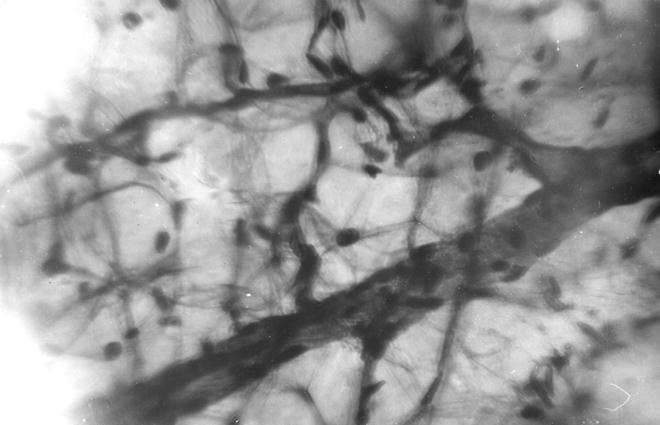
Рис. 121. Сплетение сосудов микроциркуляторного русла и вен костного мозга диафиза плечевой кости теленка новорожденного периода. Импрегнация свинцом МБИ-6 (Б. В. Криштофорова, 1974).
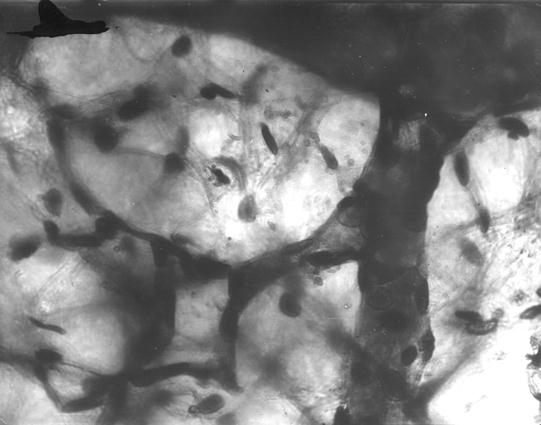
Рис. 122. Слияние венозных ветвей в диафизарную вену плечевой кости теленка (сутки). Импрегнация свинцом. МБИ-6, 10х8 (Б. В. Криштофорова, 1974).
Рис. 123. Стенка внутрикостной вены плечевой кости теленка (20 суток). Импрегнация свинцом. МБИ-6, 7х40 (Б. В. Криштофорова, 1974).
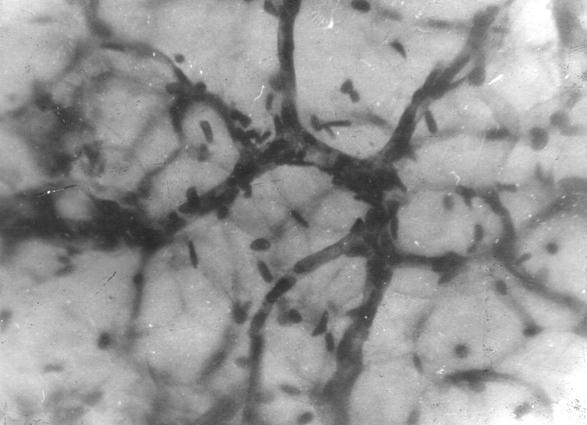
Рис. 124. Слияние внутрикостных вен бедренной кости теленка (20 суток). Импрегнация свинцом. МБИ-6, 10х20 (Б. В. Криштофорова, 1974).
.
Рис. 125. Посткапиляры и венула плечевой кости теленка (сутки). Импрегнация свинцом. МБИ-6, 10х20 (Б. В. Криштофорова, 1974).
У новорожденных (суточных) телят в коротких трубчатых костях (путовой и венечной) разветвляются кровеносные сосуды по большей части по магистральному типу. В среднем участке диафиза содержатся наибольшего калибра артерии и вены. Как и в эпифизах длинных трубчатых костей, вены образуют параартериальные сплетения. Кроме того, в диафизе путовой кости оказывается значительного размера венозный коллектор (рис. 126).
В проксимальных и дистальных участках диафиза путовых и венечных костей суточных телят количество вен больше чем артерий (табл. 159). Вокруг некоторых вен формируются паравенозные сплетения из мелких артерий и артериол, которые побуждают течение крови по этим венам [192-195].
Особенное строение имеют артерии и вены копытцевой кости (рис. 127). Терминальная артерия проникает в нее сквозь аксиальное отверстие канала подошвы, по поти, отдавая ветви разного калибра в мягкие ткани копытца. Многочисленные внутриканальные вены размещаются вокруг артериальной магистрали и образуют структуру подобную кавернозному телу (рис. 128).
Кровеносные сосуды губчатой костной ткани (энхондрального происхождение) копытцевой кости образуют подобные сплетения, как и в эпифизах трубчатых костей конечностей [193].
Структура стенок артерий, вен и сосудов микроциркуляторного русла отвечает таким же особенностям, как и в других костных органах. Однако, в копытцевой кости стенка венозных магистралей имеет свои особенности. Под слоем эндотелия содержится тонкий эласто-коллагеновый каркас, который переходит во внешний слой рыхлой волокнистой соединительной ткани, что, возможно, предопределено увеличением давления при опоре конечности. У суточных телят вены и артерии копытцевых костей имеют наибольший калибр и суммарный поперечник по сравнению с венечными и путовыми костями [193].
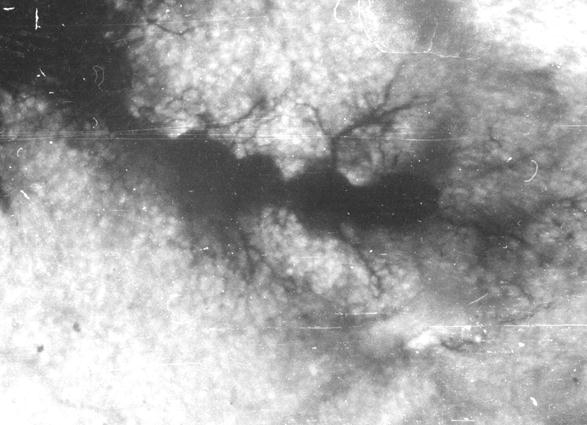
Рис. 126. Просветленный препарат путовой кости суточного теленка. Инъекция тушью с желатином, МБС-10, 8х2 (В. В. Лемещенко). 1. Четковидная вена диафиза. 2. Костный мозг.
С возрастом телят, особенно при активной локомоции, происходит значительный рост калибра артерий и вен на фоне уменьшения его в сосудах микроциркуляторного русла при замещении красного костного мозга желтым.
В основных венозных магистралях трубчатых костей новорожденных телят встречаются внутривенные образования в виде складок интима, клапанов, трабекул и артерий, которые пронизывают их просвет [133]. Возможно, что они принимают участие в направлении течения крови и предопределяют тонких стенок вен при повышении гидростатического давления.
Значительное количество анастомозов между интраоссальными сосудами обеспечивает течение крови в разных направлениях соответственно биомеханических нагрузок, которые возникают при локомоции животных. Венозные анастомозы есть не только в отдельных участках костных органов, но и между ними, что используется для проведения внутрикостных инъекций лекарственных веществ [133, 138]. Через интраоссальную сетку вен любого костного органа можно наполнить всю венозную систему организма [133].
Иннервация костных органов достаточно сложна и недостаточно исследованная, что обусловленно их минерализацией. На данное время больше освещена иннервация надкостницы, несколько меньше костного мозга и совсем мало – костной ткани. Однако необходимо отметить, что разные нервные волокна можно обнаруживать в отдельных костных трабекулах и остеонах. И. В. Хрусталева [334] отмечает, что остеорецепция не только влияет на функции костного органа, но и на весь организм. Источником иннервации костных органов являются все нервные стволы, которые проходят в окружающих кость мягких тканях. Каждый костный орган всегда получает нервы от нескольких центров нервной системы [332-334]. Практически денервировать костный орган невозможно.
|
|
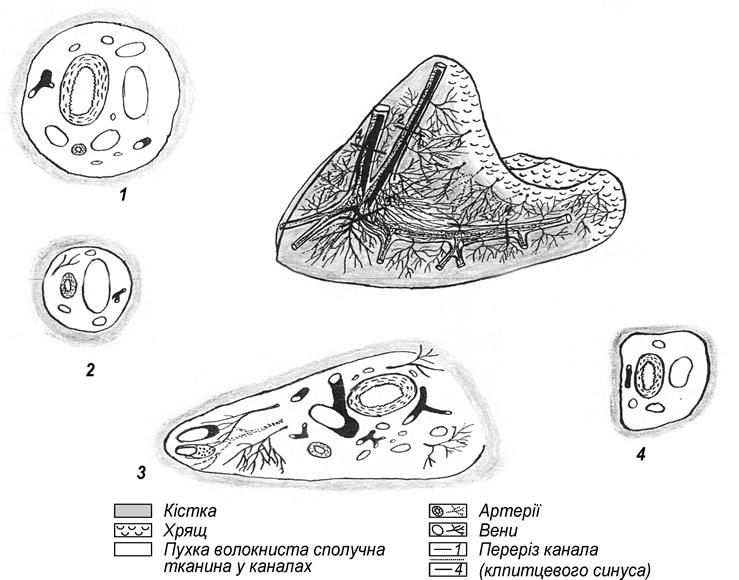
Рис. 127. Схема кровеносных сосудов копытцевой кости новорожденных телят (В. В. Лемещенко, 1996).
Артериовенозные взаимоотношения в каналах
|
№ п/п |
Поперечник артерий, мкм |
Вены* |
|
1 |
650-750 |
500-600 100-400 50-75 1 2-8 5-7 |
|
2 |
150-175 |
200-400 25-75 1 2-5 |
|
3 |
600-750 |
550-600 200-400 25-100 1 1-2 дек. дес. |
|
4 |
300-400 |
100-250 50-75 1(0) 2-5 |
*В числителе указан поперечник вен (мкм), в знаменателе их количество
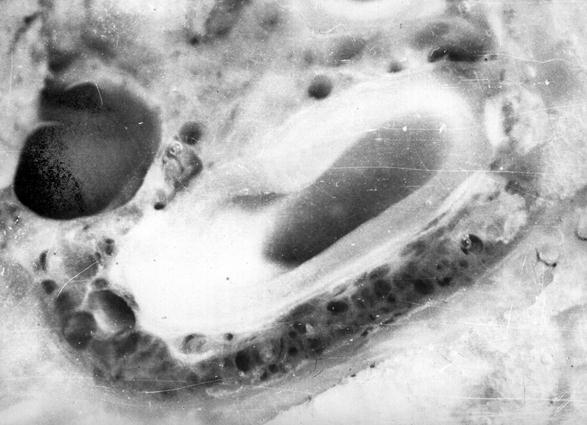
Рис. 128. Декальцинированный срез копытцевой кости 5,5-летней коровы (В. В. Лемещенко, 1997): 1 – копытцевая артерия; 2 – параартериальные вены; 3 – “Пещеристое тело”; 4 – костная балка.
Таблица 163
Относительная площадь артерий и вен губчатой костной ткани коротких трубчатых костей пальцев телочек красной степной породы, мкм (В. В. Лемещенко, 1997)
|
Возраст |
Кости |
Конечности |
Проксимальный Эпифиз |
Проксимальный участок |
Средний участок диафиза |
Дистальный участок диафиза |
|
Новорожденные |
Путовая |
Г* |
8,80±1,21 |
11,27±1,10 |
7,34±1,34 |
7,61±1,41 |
|
Т |
9,04±1,2 |
10,64±1,15 |
5,95±1,13 |
7,21±1,39 |
||
|
Венечная |
Г |
9,84±1,11 |
11,05±1,25 |
8,32±1,57 |
9,47±1,23 |
|
|
Т |
8,80±0,47 |
7,14±0,81 |
3,93±0,91 |
6,78±1,33 |
||
|
10 суточные |
Путовая |
Г |
5,64–7,34 |
11,!3–11,84 |
2,28–2,48 |
6,75–7,34 |
|
Т |
4,44–5,16 |
9,43–10,24 |
1,88–2,38 |
4,68–5,56 |
||
|
Венечная |
Т |
5,95–9,26 |
7,54–11,11 |
1,32–3,97 |
6,22–8,43 |
|
|
Г |
6,00–6,74 |
7,14–7,46 |
2,25–3,75 |
5,56–5,82 |
* Г и Т – грудные и тазовые конечности
Таблица 164
Суммарный поперечник артерий и вен костей пальцев новорожденных телят красной степной породы, мкм (В. В. Лемещенко, 1997)
|
Название |
Возраст, сутки |
Конечность |
|||
|
Грудная |
Тазовая |
||||
|
Артерии |
Вены |
Артерии |
Вены |
||
|
Путовые |
1 |
526,33±97,58 |
1896,03±356,71 |
832±50 102,43 |
2232±33 434,74 |
|
10 |
871,97–896,20 |
1827,80–2306,00 |
772,07–854,50 |
1191,00–1503,20 |
|
|
Венечные |
1 |
432,90±50,19 |
1285±60401,60 |
658,60±37,00 |
1552,15±135,05 |
|
10 |
498,27–862,10 |
964,47–1132,20 |
487,17–850,15 |
1013,80–1143,30 |
|
|
Копытцевые |
1 |
882,73±51,79 |
1356,67±263,15 |
947,20±39,27 |
1227,17±114,29 |
|
10 |
925,00–1139,60 |
1166,67–1729,13 |
810,00–1250,60 |
1324,98–2123,80 |
Суммарный поперечник сосудов определяли в среднем участке диафиза коротких трубчатых и подошвенном канале копытцевых костей.
Таким образом, костные органы – очень сложные и лабильные органы, особенно у новорожденных продуктивных животных, за счет хорошо развитой сети кровеносных сосудов и нервов.
Жизнеспособность новорожденных продуктивных животных в полной мере зависит от развития костной системы в пренатальный период онтогенеза. Одной из основных причин заболевания животных в первые сутки жизни есть нарушение процессов остеогенеза во внутриутробный период развития. У плодов с началом остеогенеза в костных органах совершается образования всех клеток крови и иммунной системы. В этот же период костная система активно включается в обмен веществ, особенно водный и минеральный [176, 177].
У новорожденных проидуктивных животных проявляются генотипически закономерности формы остеометрических параметров и интраорганных структурах костных органов в зависимости от биомеханической функции. Потому при содержании родителей в условиях гиподинамии возникает задержка костеобразования, а недостаток в рационе минеральных веществ приводит к образованию хрящевых трабекул остеоида. Это одна из главных причин, которая предопределяет нарушения роста и развития костной системы, которая отмечается у неонатальных продуктивных животных.