Система скелетных мышц
Одной из наиболее динамических систем аппарата движения есть система скелетных мышц. Их сокращение предопределяет эффект движения в пространстве и времени всего организма или его отдельных частей, а также обеспечивает изменение объема полостей тела. Функция респираторных мышц (инспираторов и экспираторов) включается в работу с первых минут после рождения животного, и как сердце ритмично работает обеспечивая жизнь. Она является основным проявлением жизни организма до последних минут его жизни. Мышцы живота предопределяют уменьшение или увеличение объема брюшной полости.
Система скелетных мышц присуща лишь высшим позвоночным (в отличие от всех других животных), она построена из поперечно - исчерчененных мышц и дифференцирована на более 200 мышц как органов, связанных с центрами мозга непрерывной и очень быстрой связью – это целая система органов, которые способны к сокращению и этим самым вызывают эффект движения. Закрепляясь на костных органах, как на системе рычагов, мышцы, сокращаясь, образуют разные виды движения и перемещения организма в пространстве, закрепляют костные органы в определенном соединении, храня форму тела и его положения.
Большое значение система скелетных мышц имеет в проявлении жизнеспособности организма у матуронатных животных. У телят через 20-30 минут после рождения нарастает тонус мышц, который обеспечивает проявление генетической доминанты не только статики, но и движения в пространстве и времени, которое определяется как локомоция.
Генетическая детерминированность размещения скелетных мышц на туловище, грудных и тазовых конечностях подтверждается наличием коррелятивных связей (табл. 165). Связь между живой массой суточных телят и массой скелетных мышц, которые расположены на туловище тесный и позитивный (r=0,78), но несколько меньше чем с массой скелетных мышц грудных конечностей (r=0,83) и, особенно, тазовых (r=0,92). Характерно, что у телочек красной степной породы, полученных от коров выращиваемых с использованием принудительного дозированного движения, эти взаимосвязи очень тесны и позитивны – коэффициент корреляции колеблется в пределах r=0,94-0,99. У пренотальных недоразвитых телочек они также очень тесные и позитивные (r=0,99), что свидетельствует о продолжении роста развития плода на позднем этапе гестации. С возрастом, у 10-суточных телочек, взаимосвязи между живой массой и массой скелетных мышц оказываются как средние и обратнозависимые (r=0,57). Они несколько растут между живой массой и массой скелетных мышц грудными конечностями (r=-0,78), а с тазовыми – почти исчезают. У пренотальных недоразвитых телочек связки становятся обратными и могут исчезать совсем. Это свидетельствует о значительном снижении биомеханических нагрузок на аппарат движения у пренотальных недоразвитых животных [3].
Таблица 165
Коррелятивные взаимосвязи между живой массой и массой скелетных мышц неонатальных телят, С. О. Акулов, 1997)
|
Возраст, суток |
Масса |
Масса скелетных мышц туловища |
Масса скелетных мышц грудных |
Масса скелетных мышц тазовых |
|
Суточные (гиподинамия) |
0,94 |
0,78 |
0,83 |
0,92 |
|
Суточные (ДПД) |
0,98 |
0,99 |
0,96 |
0,94 |
|
Суточные (недоразвитые) |
0,99 |
0,99 |
0,99 |
0,99 |
|
10-суточные |
-0,23 |
-0,21 |
-0,50 |
0,09 |
|
Между массой скелета и массой скелетных мышц |
||||
|
Суточные (гиподинамия) |
0,98 |
0,68 |
0,93 |
0,92 |
|
Суточные (ДПД) |
0,96 |
0,98 |
0,95 |
0,89 |
|
Суточные (недоразвитые) |
0,99 |
0,99 |
0,98 |
0,99 |
|
10 суточные |
-0,57 |
-0,43 |
-0,78 |
-0,13 |
* ДПД – дозировано принудительное движение
Большая потеря энергии при рождении вызывает чувство голода, который проявляется поиском источника корма. Первые шаги – это первое проявление функции аппарата движения у новорожденных животных.
Рост, развитие и формирование скелетных мышц особенно интенсивно проходит в конце пренатального периода онтогенеза. После рождения на скелетные мышцы влияют два фактора: силы гравитации и масса организма. Следует отметить, что в наибольшей работе мышц нуждается статика, как всего организма, так и его частей, что предопределяет функцию каждой мышцы определяет его структуру, форму и силу.
У новорожденных телят скелетные мышцы, как органы локомоторного аппарата, характеризуются общей структурой рядом с отличиями каждой мышцы, в зависимости от места его локализации на скелете. При световой микроскопии мышцы новорожденных животных характеризуются незавершенностью структур. Во всех мышцах выраженная продольная исчерченность, которая предопределена особенным размещением миофибрилл в мускульном волокне. Поперечная исчерченность, какая присущая скелетной мускульной ткани, определяется лишь в отдельных участках и имеет лестничный характер, который свидетельствует о рыхлом расположении сократительных элементов мышечного волокна [3].
Среди мускульных волокон встречаются дегенеративные, в ядрах которых содержится незначительное количество хроматина. В участках, где происходит непосредственное разрушение мышечного волокна, не определяется и продольная полосатость. Дегенерирующие мышечные волокна больше всего оказываются в экстензорах тазовых конечностей, что свидетельствует об интенсивной их перестройке соответственно действию силам натяжения.
Ядра мышечных волокон у новорожденных телят, расположенные преимущественно на периферии под сарколеммой между фибриллами. Как исключение, некоторые ядра могут быть одиночные или парами располагаясь в центре мышечного волокна с направлением заостренных концов под углом к сарколемме. По структуре ядра мышечных волокон можно разделить на три вида. Первый вид – это округлые ядра, бедные хроматином и имеют 1-2 ядрышка. Второй вид – продолговатые ядра, богатые хроматином и содержат 5-7 ядрышек. К третьему виду (бывают редко) относятся ядра также богатые хроматином, но имеют лишь 1-3 ядрышка [3].
Площадь мышечных волокон колеблется от 173,11 до 401,30 мм2. На фронтальных срезах мышечные волокна скелетных мышц новорожденных телочек имеют полигональную или округлую форму. Характерно, что округлые мышечные волокна окружены полигональными. Прослойки рыхлой волокнистой соединительной ткани между отдельными мышечными волокнами составляют от 11,99±1,75 до 56,29±4,68 % на гистотопограмме. Поперечник мышечного волокна колеблется от 14,85±0,85 до 21,61±1,50 мкм.
Доказано, что у новорожденных телят коэффициент подражания мясной производительности 0,11-0,53, нежности мышц – 0,60-0,71, размещение и нахождение жира – 0,09 [269].
У новорожденных статическая функция в первые часы и сутки после рождения недостаточная. Они, пытаясь встать на конечности, часто падают, спотыкаются. Статическая функция мышц с их сухожилиями проявляется у телят, жеребят до конца первых суток, поросят – на 4-6 сутки, а щенков – на 12-16.
Масса скелетных мышц в постнатальном периоде онтогенеза новорожденных животных характеризуется неравномерностью ее роста [205]. Мышцы теленка при рождении образуют механизм, через какой организм теряет органические запасы. Движение необходимо для пополнения этих запасов, а это значит и для существования. Относительная масса отдельных мышц, в этой сложной системе, определенная также эволюционным наследством вида, которая предназначена для пополнения прямой потребности новорожденного, а затем в процессе роста в постнатальном периоде онтогенеза, рядом с увеличением общей массы. Рост каждой мышцы детерминирован генетически и факторами окружающей среды, которые влияют, на фоне эволюционного развития вида, и определяют закономерность ее периодичности, а также в ответ на повседневную активность животного [43].
У новорожденных телочек, полученных от коров стойлово-привязанной технологии содержания, живой массой 35,70±2,32 кг абсолютная масса скелетных мышц достигает 12,54±0,76 кг или же 35,13% относительной массы. Характерно, что масса костных органов в два раза меньше (7,25±0,51 кг) или 20,34% [3].
Необходимо отметить, что использование при стойлово-привязанной технологии содержания дозированного принудительного движения продуктивных животных при равнозначной живой массе (35,22±2.87 кг) приводит к росту абсолютной массы скелетных мышц (15,11±1,09 кг), как и относительной (42,90%). Можно допустить, что только механическая энергия, которая возникает при движении стельных коров, предопределяет рост массы скелетных мышц на новорожденного7,77% и побуждая оптимальный рост и развитие его аппарата движения, и проявляясь соответствующими экстерьерными характеристиками. Масса скелетных органов при этом увеличивается до 21,98% (табл. 166). Это дает основание предполагать, что пренатальное развитие костных органов больше определяется генетически чем мышц. То должно учитываться при откорме: масса скелетных мышц аппарата движения новорожденных телят, полученных от стельных коров выращиваемых в условиях гиподинамии, на 38,58% меньше, чем тех, которые выращивали с использованием дозированного принудительного движения.
Таблица 166
Морфометрические особенности скелетных мышц неонатальных телят (С. О. Акулов, 1997)
|
Возраст |
Живая масса, кг |
Масса скелета |
Масса скелетных мышц |
||
|
Абсол., кг |
Относ., % |
Абсол., кг |
Относ., % |
||
|
Суточные (гиподинамия) |
35,70±2,32 |
7,26±0,51 |
20,34 |
12,54±0,76 |
35,13 |
|
Суточные (дозировано принудительное движение) |
35,22±2,87 |
7,39±0,59 |
21,98 |
15,11±1,09 |
42,90 |
|
Суточные (недоразвитые) |
29,83±1,17 |
7,18±0,47 |
31,11 |
9,28±0,91 |
31,11 |
|
10-суточные |
40,70±2,63 |
7,93±0,27 |
19,48 |
13,70±±0,92 |
33,66 |
Анализ абсолютной и относительной массы отдельных скелетных мышц размещенных на определенных участках скелета показывает, что она наибольшая на туловище (табл. 167). У телочек, которые родились от коров при содержании в условиях гиподинамии, составляет 40,09% живой массы. У телочек, полученных от коров, которые получали дозовое принудительное движение, – значительно больше – 48,60% (на 4,51%), а в недоразвитых – наименьшая 33,48% (на 0,61%). Относительная их масса, по сравнению с массой скелета, составляет 69,28%, 88,65 и 56,00% (соответственно). Такая же закономерность в изменении относительной массы скелетных мышц отмечается и на конечностях новорожденных телочек. Однако наибольшая относительная масса мышц регистрируется на тазовых конечностях. Относительно живой массы они составляют 13,05%,14,91 и 13,07% (соответственно). Степень роста массы скелетных мышц в пренатальный период онтогенеза в зависимости от двигательной активности коров еще больше оказывается по отношению ее к массе скелета [3]. У недоразвитых телочек она меньше на 43,50-75,24%. Масса скелетных мышц грудных конечностей телочек где-то на 6% более малая от тазовых. Использование дозового принудительного движения коровам также способствует росту скелетных мышц в пренатальный период онтогенеза. Пренатальное недоразвитие новорожденных телочек характеризуется уменьшением массы скелетных мышц конечностей.
Таблица 167
Абсолютная и относительная масса скелетных мышц туловища и конечностей неонатальных телят (С. О. Акулов, 1997)
|
Возраст |
Абсолютная масса, кг |
Относительная масса % к массе тела |
||||
|
М.* Туловища |
М. Грудных К.* |
М. Тазовых К. |
М. Туловища |
М. |
М. |
|
|
Суточные (гиподинамия) |
5,03±0,15 |
2,86±0,23 |
4,66±0,45 |
14,09 |
8,01 |
13,05 |
|
Суточные (ДПД) |
6,55±0,43 |
3,31±0,34 |
5,25±0,34 |
11,60 |
8,40 |
14,91 |
|
Суточные (недоразвитые) |
4,02±0,17 |
2,01±6,29 |
3,90±0,39 |
13,48 |
6,97 |
13,07 |
|
10-суточные |
5,91±0,41 |
2,84±0,18 |
4,96±0,37 |
14,52 |
6,98 |
12,19 |
М*– мышцы, К* – конечности
У новорожденных телочек отдельные группы скелетных мышц имеют разную интенсивность пренатального роста и развития. Длиннейшая мышца груди и поясницы, у телочек рожденных от коров при технологии содержания с использованием дозированного принудительного движения достигает 288,40±24,59 г, тогда как при гиподинамии – 273,31±21,25 г, а у недоразвитых – 197,20–13,26 г (табл. 168).
Таблица 168
Морфометрические характеристики мускульных волокон (МВ) длинейшей мышцы груди и поясницы (С. О. Акулов, 1997)
|
Возраст |
Живая |
Поперечник, мкм |
Количество МВ |
Средняя |
% рыхлой соединительной |
|
Суточные (гиподинамия) |
35,70±2,32 |
21,14±1,91 |
1900,00±5,00 |
350,82 |
12,23±1,01 |
|
Суточные (ДПД) |
35,22±2,87 |
22,03±1,33 |
1870,09±11,23 |
380,98 |
12,34±2,01 |
|
Суточные (недоразвитые) |
29,83±1,17 |
20,97±1,74 |
1893,20±30,07 |
345,20 |
13,61±1,81 |
|
10-суточные |
40,70–42,49 |
20,55–1,06 |
1846,67±6,02 |
331,50 |
12,00±1,47 |
Таблица 169
Динамика абсолютной и относительной массы некоторых скелетных мышц к общей массе мускульной кости телочек (Акулов С. О., 1997)
|
Возраст, |
Масса |
Мышцы |
||||
|
Длиннейшей мышцы груди |
Двуглавая |
Трехглавая |
Четырехглавая Бедра |
Полусухожильная |
||
|
1 |
Абсолютная, г |
197,20-288,40 |
33,30-42,06 |
215,32-289,00 |
330,65-429,32 |
112,48-129,14 |
|
Относительная % |
1,91-2,18 |
0,28-0,36 |
1,91-2,32 |
2,81-3,56 |
0,80-1,21 |
|
|
10 |
Абсолютная, г |
359,96-365,87 |
47,32-52,44 |
292,55-304,10 |
433,76-517,33 |
157,33-169,98 |
|
Относительная % |
4,14-4,54 |
0,59-0,60 |
3,44-3,69 |
5,47-5,85 |
1,92-1,92 |
На грудных конечностях наблюдается подобная закономерность: трехглавая мышца плеча имеет почти такую же абсолютную массу, как и длиннейшая мышца груди и поясницы (табл. 169).
У новорожденных телочек, абсолютная масса двуглавой мышцы плеча, которая действует на плечевой и локтевой суставы, наименьшая [3]. Однако она опять же наибольшая у телочек, рожденных от коров, которые получали дозированное принудительное движение. На тазовой конечности наибольшая абсолютная масса отмечается в четырехглавой мышце бедра (429,32±20,67 г), тогда как у недоразвитых телочек она значительно снижается (330, 65±16,69 г).
Необходимо отметить, что к рождению продуктивных животных каждая скелетная мышца имеет определенную структуру, что, возможно, предопределенно разной интенсивностью их роста и развития и генетически детерминировано (рис. 129, 130). Мышечные волокна имеют почти одинаковый поперечник. Однако, количество мышечных волокон у пренотальных развитых суточных телят значительно больше, а прослоек соединительной ткани между ними на 1-2% меньше.
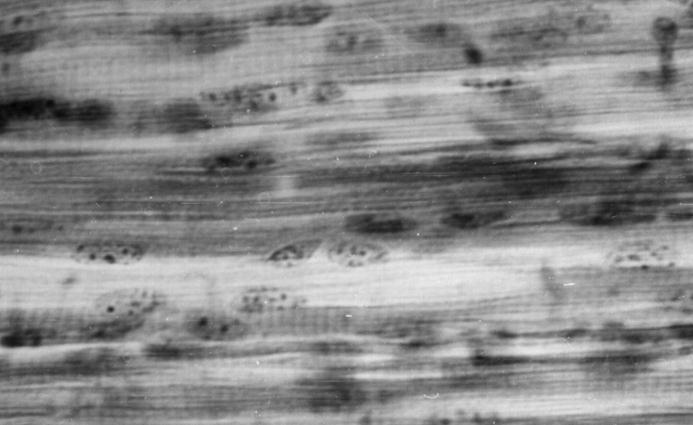
Рис. 129. Мышечные волокна длиннейшей мышцы груди и поясницы теленка (сутки). Гематоксилин и эозин, МБИ-6, х400 (С. О. Акулов, 1998).

Рис. 130. Мышечные волокна длиннейшей мышцы груди и поясницы теленка (10-суток). Гематоксилин и эозин. МБИ-6, х400 (С. О. Акулов, 1998).
Состояние мышц на скелете новорожденных животных неравномерно, но наиболее они развиты в участке бедра, несколлько меньше – лопатки, крупа и позвоночника и менее всего в среднем и дистальном звеньях конечностей.
В мышцах новорожденных телят содержится воды – 81,9%, сухих веществ – 18,1%. В первые же сутки жизни количество воды в мышцах уменьшается до 80,57%, а сухих веществ, напротив, увеличивается до 19,43%. Характерно, что сухие вещества состоят из белков и совсем незначительного количества жира. Состав скелетных мышц поросят подан в таблицах 170 и 171. У суточных поросят очень низкое содержание белковых фракций (11,0г/кг), количество которых на конец новорожденного периода вырастает больше в 2 раза (25,6 г/кг). В мышечных волокнах содержится значительное количество белка фибрилл.
Таблица 170.Азотные фракции скелетных мышц поросят, г/кг
|
Показатели |
Возраст, суток |
|
|
1 |
21 |
|
|
Небелковый |
2,6 |
3,5 |
|
Белковый |
11,0 |
25,6 |
|
Белок саркоплазмы |
2,2 |
5,4 |
|
Белок фибрилл |
6,5 |
16,4 |
|
Внеклеточный белок |
2,3 |
3,2 |
|
Коллагеновый белок |
1,3 |
3,0 |
Таблица 171
Минеральный состав скелетных мышц поросят (1кг)
|
Показатели |
Возраст, суток |
|
|
1 |
21 |
|
|
Живая масса, кг |
1,21 |
3,45 |
|
Вода % |
82,3 |
77,8 |
|
Общий азот, г |
15,4 |
29,1 |
|
Na, мекв |
54,0 |
34,0 |
|
K, мекв |
73,0 |
102 |
|
Cl, мекв |
37,0 |
32 |
|
P, ммоль |
57,0 |
77 |
|
Mg, мекв |
15,3 |
16,2 |
|
Cu, мекв |
2,9 |
3,3 |
Воды в скелетных мышцах суточных поросят содержится 82,3%, а 21-суточных – 77,8. Количество общего азота достигает 15,4г и 29,1г. В скелетных мышцах оказывается определенное количество макро - и микроэлементов.
Избыток влаги в мышцах новорожденных животных предопределяет округлость форм участков тела, особенно крупа и бедра. Однако на разрезе таких мышц влага свободно выступает на поверхность, которая указывает на нарушение водного обмена у животных низкой жизнеспособности еще до рождения. Необходимо помнить, что у таких животных кожный покров легко отделяется от ниже расположенных тканей. И наоборот, когда новорожденное животное погибает от обезвоживания кожный покров снимается с определенным усилием, а подкожная основа и рыхлая волокнистая соединительная ткань имеет матовый и белесоватость цвет, тогда как мышечные волокна на поперечном разрезе выделяются в виде отдельных пучков.
Необходимо отметить, что подобных сведений об особенностях строения скелетных мышц у других видов новорожденных животных мы не обнаружили.
Таким образом, у новорожденных матуронатных животных, особенно у телят и жеребят аппарат движения способен выполнять определенные биомеханические функции – статику и динамику. Это дает возможность выполнять как двигательные акты, так и проявлять на определенном уровне рефлекс сосания. При определенной двигательной активности новорожденные животные получают биомеханическую энергию и лучше растут и развиваются. Двигательная активность матуронатных новорожденных в первые сутки жизни имеет определенные проявления, которые чередуются статикой и локомоций.
Исследования А. А. Бачманова [21] показывают, что у новорожденных телят заключенных в клетки-пеналы двигательная активность на второй день жизни несколько снижается, а на третий – опять растет (табл. 172). Возможно, это предопределено недоразвитостью аппарата движения, а также условиями в которых находится теленок (узкие клетки). На 5 сутки двигательная активность свободно двигающихся телят достигает 22,4% всего времени, а частота голосовых актов растет до 183,5%.
Таблица 172
Этологическая характеристика телят в первые сутки жизни (А. А. Бачманов, 1987)
|
Возраст, |
Длительность |
Подвижная |
Частота голосовых актов |
|||
|
Минут |
% |
Минут |
% |
Минут |
% |
|
|
1 |
25,8±1,9 |
61,1 |
3,7±0,1 |
33,4 |
1,5±0,2 |
111,5 |
|
2 |
12,6±0,9 |
61,1 |
3,7±0,1 |
25,9 |
0,6±0,1 |
135,8 |
|
3 |
16,3±1,0 |
47,0 |
3,6±0,1 |
20,5 |
0,9±0,2 |
150,7 |
|
4 |
16,7±2,5 |
65,0 |
3,7±0,1 |
24,0 |
0,5±0,1 |
183,3 |
|
5 |
15,2±1,6 |
51,2 |
3,7±0,2 |
22,4 |
0,4±0,1 |
183,5 |
Следовательно, аппарат движения зрелорождающихся животных содержащихся вне клеток имеет определенную незавершенность строения, однако из первых минут после рождения обеспечивает выполнение двигательных актов. В пренатальном развитии аппарата движения особенное значение имеет остеогенез, который обеспечивает не только подвижность животного, но и обеспечивает определенное становление защитных функций организма и гемоиммунопоэза. У незрелорождающихся животных в пренатальный период преимущественно формируются только диафизарные центры окостенения. Хрящевые эпифизы характерны не только для костных органов осевого скелету, но и для скелета конечностей.
У пренатально недоразвитых матуронатных и имматуронатных новорожденных отмечается нарушение остеогенеза, который приводит к снижению не только биомеханической функции костной системы, но и гемоиммунопоэза, уже предопределяя снижение жизнеспособности животных.