СИМФЕРОПОЛЬ - 2009 УДК 619: 577.27
Рекомендации разработаны на основе результатов научно-исследовательской работы «исследовать особенности морфогенеза иммунокомпетентных структур и разработать критерии морфологической оценки их недоразвития во взаимосвязи с жизнеспособностью животных неонатального периода».
Предложены рекомендации по технологии выращивания телят с момента рождения до конца новорожденного периода, который совпадает с началом функционирования рубца. При этом принимаются во внимание особенности пренатального развития новорожденных телят по разработанным тестовым показателям. В основу определения тестовых показателей пренатального развития телят положены морфофункциональные особенности их органов универсального гемоиммунопоэза и лимфоцитопоэза. Рекомендации позволяют повысить не только сохранность телят, но и обусловливают полную реализацию генетических потенций их роста и развития и жизнеспособности в новорожденный и молочный периоды. Также в них поданы меры, предотвращающие заболевание с проявлениями нарушения функции органов пищеварения у телят.
Приведено полное описание морфофункциональных особенностей органов гемоимунопоэза и методических подходов по выяснению и коррекции организменного статуса домашних животных. Данные предоставлены в форме, которая максимально способствует их практическому использованию. Рекомендации предназначены для врачей ветеринарной медицины, владельцев скотоводческих хозяйств и технологов животноводства.
Под редакцией Б. В. Криштофоровой.
Разработчики: сотрудники кафедры анатомии и физиологии животных Южного филиала Национального университета биоресурсов и природопользования Украины»Крымский агротехнологический университет"
Б. В. Криштофорова, доктор ветеринарных наук, профессор;
В. В. Лемещенко, доктор ветеринарных наук, профессор;
В. Г. Соколов, кандидат ветеринарных наук, доцент.
Рецензенты: В. Т. Хомич, доктор ветеринарных наук, профессор;
В. Л. Ковалев, доктор ветеринарных наук, профессор.
ВСТУПЛЕНИЕ
Проблемы выращивания здоровых продуктивных животных в настоящее время имеют общие черты для всех стран мира, независимо от форм хозяйствования. И это потому, что они обусловлены возрастающим действием антропогенных факторов и технизацией производственных процессов. Определенная изоляция животных от условий окружающей среды, рост во внешней среде метаболитов человеческой деятельности особенно негативно отражаются на защитных свойствах их организма, вызывая не только нарушения роста и развития, но и способствуют распространению различных заболеваний.
Проявлением снижения иммунного статуса животных в условиях их содержания, прежде всего, стало рождение недоразвитого приплода. Переболевшие животные отстают в росте и развитии, не реализуют генетических потенций продуктивности, дают еще более слабый приплод, приводя к замыканию “порочного круга”.
При отсутствии природных факторов, особенно в период новорожденности, не полностью реализуется запрограммированный в организме генетический потенциал его роста и развития. Как результат, снижаются адаптивные возможности и жизнеспособность организма животных, что проявляется сокращением продолжительности их биологической жизни. Часто коров используют, в основном, не более 2-3 отелей.
Признаки деградации вида проявляются, прежде всего, в прогрессивном снижении функции воспроизводства. Оплодотворение самок продуктивных животных достигает 60-70%, птицы – 25-60%. На 100 голов коров в хозяйствах получают 70-80 телят при биологической возможности 100-105. увеличивается продолжительность сервис-периода, возрастает количество случаев внутриутробной гибели плодов, патологических родов и послеродовых осложнений. Все это свидетельствует о том, что в будущем может исчезнуть определенный вид животных, как это происходит в естественном биоценозе.
В практической работе врачу ветеринарной медицины и животноводам очень часто приходится принимать различные меры с целью сохранения новорожденных животных. Сейчас значительное загрязнение окружающей среды, которое является следствием технизации производственных процессов и факторов человеческой деятельности, обусловило изменения в отношениях в системе «мать-плацента-плод» рождения только 5-7% продуктивных животных, статус организма которых соответствует гестации. Они, по экстерьерным характеристикам, проявлению двигательной активности, рефлексу сосания и другим безусловным рефлексам соответствуют породным показателям, установленным для них еще в XVIII-XIX веках.
Общепринятая технология при таком условии не требовала пристального надзора за новорожденными, а только заблаговременного кормления молозивом матери и содержания в соответствующих помещениях. Сейчас новорожденных телят размещают в отдельных клетках сразу после рождения, кормят молозивом по общепринятой норме – 2 литра. При этом ни врач ветеринарной медицины, ни доярки (телятницы) не обращают внимание на морфофункциональное состояние организма новорожденного. В гораздо больших случаях не принимают во внимание время рождения теленка, которое от момента отела до кормления колеблется от 30 минут до 10 часов и более. Конечно, при рождении теленок расходует большое количество энергии, которая нуждается в пополнении за счет молозива. Значительное количество иммуноглобулинов в молозиве в таком случае не являются стартовыми, которые побуждают функциональную активность желез тонкой кишки и являются запороговым действующим фактором на иммунные образования, содержащиеся в нем. В таком случае первое кормление вызывает нарушение пищеварения, проявляясь выделением жидких фекалий, окраска которых почти совпадает с таковым молозива. По условию своевременного принятия мер, не требующих значительных затрат (замена молозива настоями и отварами трав), такое нарушение проходит через 1-2 суток без заметных изменений. Однако даже в случае рождения теленка морфофункциональный статус организма которых совпадает с породными показателями, первые признаки нарушения функции органов пищеварения не принимаются во внимание обслуживающим персоналом. Только в случае, когда теленок отказывается от приема корма, а состояние его организма значительно ухудшилось, сообщают о болезни врачу ветеринарной медицины. Осмотрев больного теленка, врач принимает соответствующие лечебные меры, что дает возможность спасти жизнь животного, однако, уже происходит нарушение роста и развития организма. Суточный прирост живой массы становится минимальным и не превышает 70 г, что приводит к росту экономических затрат. На этом фоне растет экономические затраты.
Более негативные результаты получают животноводы по условию рождения пренатально недоразвитого теленка с резкой проявлением клинических признаков.
Этологические исследования свидетельствуют, что по условию не достаточно весомого пренатального недоразвития у новорожденных телят происходит только несколько незначительная задержка доминант: двигательной активности и проявления рефлекса сосания (до 45-60 мин.). Однако значительное недоразвитие новорожденных телят, которое проявляется уже наглядно-уменьшением живой массы, промеров полов тела, красноватой окраской носогубного зеркала, отсутствием резцовых зубов на нижней челюсти свидетельствует о том, что для сохранения жизни такому теленку необходимо особое внимание к условиям его кормления и выращивания. Пренатально недоразвитых телят в настоящее время рождается более 60%. Как свидетельствуют наши исследования, фактором, который не только подавляет рост и развитие иммунных образований, но и вызывает повышенную их чувственность к иммуноглобулинам молозива является нарушение плацентарного барьера.
В таком случае кормление новорожденных телят молозивом, особенно запоздалое, вызывает нарушения кишечного типа пищеварения На фоне быстрого развития аллергии местной или общей реакции. В таком случае кормление телят молозивом бывает первой и последней. Такое состояние возникает потому, что молозиво в меньшей степени действует как фактор восполнения энергетических затрат, но способствует повышению перистальтики кишечника и проникновению в кровообращение токсинов. Этому способствует особая функция печени, которая присуща для всех новорожденных млекопитающих. Во-первых, паренхима печени имеет незавершенное строение, которое негативно влияет на реализацию ее антитоксической функции. Кроме того, наличие венозного протока, который соединяет пупочно-воротный коллектор (образованный остатком пупочной и воротной венами печени) с каудальной полой веной, способствуя распространению токсинов с течением крови, вызывая его общую интоксикацию. Кроме того, печень у недоразвитого теленка в неполной степени выполняет гемодинамическую функцию регуляции объема тока крови к сердцу, что в свою очередь на первом этапе приводит к «утомлению» миокарда от его чрезмерной нагрузки и повышению вязкости крови (по условию дегидратации организма), а затем к необратимым изменениям в сердечной мышце, что обуславливает миокардиодистрофию. Как следствие этого наступает гибель теленка, на фоне незначительных изменений в слизистой оболочке кишечника. Однако происходит дилатация камер сердца и миокарда, что свидетельствует о недостаточности сердечной деятельности и застойных явлениях в малом круге кровообращения. Возникает отек легких, который присущ телятам значительно старшего возраста.
Следовательно, состояние экологии не только негативно влияет на мать, но и на развитие плода, вызывая пренатальную недоразвитость. В таком случае необходимо немедленно объединить все усилия на двух направлениях:
1. создать технологические условия кормления и содержания новорожденных животных, соответствующие их биологическим потребностям.
2. остановить загрязнение экологии, а затем значительно ее улучшить.
В рекомендациях, которые мы предлагаем, наибольшее внимание выделяется первому направлению-созданию биологически обоснованных технологических приемов кормления и содержания с целью повышения жизнеспособности телят.
1. Общие положения
Для определения особенностей пренатального периода онтогенеза новорожденных продуктивных животных определенное теоретическое и практическое значение имеют общебиологические критерии статуса их организма, такие как зрелорожденность (матуронатность) и незрелорожденность (иматуронатность). Большинство продуктивных новорожденных животных относят к зрелонарожающим.
Зрелые животные, прежде всего, характеризуются способностью к относительно самостоятельному существованию с первых минут после рождения при условии наличия источника корма.
Матуронатные животные рождаются зрячими, с открытыми слуховыми отверстиями и проявлением статолокомоторных актов (вставания, фиксации тела в стоячем положении) через 15-20 минут и возможностью передвижения в пространстве с помощью конечностей. Одновременно проявляются доминанты не только на необходимость потребления корма, но и на двигательную активность.
Сейчас, в течение первых суток жизни матуронатные животные проявляют условный рефлекс на источник корма и распознают свою “кормилицу”. Проявление безусловных рефлексов у зрелорождающих животных возникает через несколько минут после рождения. Такая активность реализации безусловных рефлексов определена эволюционно, потому что новорожденные матуронатные животные с первых минут жизни находятся в условиях стада, которое быстро передвигается в пространстве и претерпевает значительные изменения среды. Кроме того, перемещение стада в пространстве вызывает значительное изменение окружающей среды, что требует более развитых органов иммунной защиты, характерных для зрелых животных.
Иматуронатные животные рождаются слепые, с закрытыми слуховыми отверстиями, локомоторные движения их проявляются только в виде ползания. Конечности при ползании только помогают протягивать туловище в пространстве. Для их существования необходима определенная забота родителей, которые не только кормят, но и ухаживают и обеспечивают их гигиену. Неспособность выполнять локомоторные акты матуронатными животными связана с особенностями их пренатального остеогенеза. Почти во всех костных органах, в том числе и трубчатых костях конечностей, при рождении обнаруживаются только диафизарные (основные) центры окостенения.
Общебиологической особенностью организма всех новорожденных (матуронатных и иматуронатных) животных является незавершенность структур, что наряду с его уязвимостью и незащищенностью, является большим достоянием возможностей быстрых адаптивных изменений в соответствии с условиями внешней среды и значительной интенсивности реализации генетических потенций формообразования.
Известно, что после рождения, матуронатные и иматуронатные животные определенное время сохраняют особенности структур позднего плодового периода. На наш взгляд, организм зрелорождающих и незрелорождающих видов животных больше образован из провизорных органов, тканей и клеток. Однако одни провизорные образования исчезают или трансформируются сравнительно быстро (мгновенно), а другие – в течение часов, суток новорожденного или даже молочного периодов. У зрелых видов животных большинство провизорных структур исчезают значительно быстрее. Телята к концу суток уже реализуют значительное количество доминант. Они приобретают уверенные и координированные движения в пространстве, теряют эмбриональные мякиши на ратичках. При этом аппарат движения таких новорожденных животных достигает 70,0-80,0%, а относительная масса костной системы – 19,5-21,5%. Характерно, что у зрелорождающих животных значительно больше развита система гемоимунопоэза.
Иматуронатные животные только на 20-21 сутки теряют особенности организма, которые присущи плодам. Органы зрения и слуха начинают функционировать только после 10-14 суток и развивается способность передвижения в пространстве на конечностях. Однако локомоторные акты довольно неуверенны, движения некоординированы, ориентация в пространстве незначительна, несколько сходна с аналогичными у матуронатных животных.
Особые структурные изменения претерпевают тканевые и клеточные структуры организма животных в новорожденный период. Матуронатные животные в течение 12-14 суток претерпевают трансформации пренатальных структур в функциональные, характерные для новорожденного периода. Доказательством такого быстрого изменения структуры органов гемоимунопоэза является интенсивная динамика морфологического и биологического составов крови. У незрелорождающих животных подобная трансформация пренатальных структур проходит в течение 20-21 суток.
Итак, общая характеристика новорожденных (суточных) животных свидетельствует, что строение их организма незавершено на разных уровнях структурной организации. Такая особенность предопределяет, с одной стороны, высокую незащищенность организма от различных факторов окружающей среды, а с другой – наибольшую возможность адаптации к этим условиям. Однако необходимо помнить и тот факт, что незавершенность структур организма новорожденных животных является одним из факторов, обусловливающих заболевания их в первые часы после рождения в течение которых проходит интенсивное разрушение пренатальных структур и образование новых.
Общебиологическое свойство структурно-функционального незавершения организма продуктивных новорожденных животных не следует сочетать с пренатальным недоразвитием и несоответствием физиологических проявлений их календарному возрасту.
Пренатальное недоразвитие новорожденных животных, особенно клинического направления, многие исследователи определяют как гипотрофию, принимая во внимание, как главный признак, лишь живую массу. Однако уменьшение живой массы новорожденных животных, регистрируемое сейчас почти во всех животноводческих хозяйствах, не является достаточно информативным показателем пренатального недоразвития новорожденного животного. Доказательством такого несоответствующего определения пренатального развития является, во-первых, нарушение как количественных, так и качественных преобразований, о чем свидетельствуют значительные и глубокие исследования новорожденных детей, проведенные в человеческой медицине. В зависимости от пренатального развития, детей разделяют на доношенных и недоношенных (даже при условии соответствия гестации). Морфофункциональный статус новорожденных детей определяют по шкале Апгара. Попытка ветеринарных акушеров адаптировать шкалу Апгара для оценки морфофункционального статуса новорожденных продуктивных животных (зрелых) практически невозможна. Недоразвитые новорожденные животные характеризуются живой массой, которая может, или не может соответствовать породным показателям. При недоразвитии в пренатальный период онтогенеза проходит задержка не только количественных, но и качественных изменений. И. А. Аршавский доказывает, что определение структурно-функциональной незрелости термином “гипотрофия" не соответствует действительности, так как недоразвитость не связана лишь с алиментарной недостаточностью. Возможно, именно поэтому неонатологи гуманной медицины больше учитывают недостаточные качественные превращения у детей в пренатальный период онтогенеза и определяют его соответственно морфофункциональному состоянию организма. Следствием такого определения является необходимость в кормлении материнским молозивом или его заменителями в определенных условиях. В ветеринарной неонатологии на новорожденных (суточных) продуктивных животных обращают внимание только в случае их заболевания, что приводит к глубоким изменениям в организме и увеличению затрат на их лечение.
На наш взгляд, пренатальная недоразвитость неонатальных животных определяется как количественными (гипотрофическими), так и качественными (гипопластическими) изменениями, которые в совокупности вызывают несоответствие структуры их организма гестации. У недоразвитых животных в пренатальном периоде онтогенеза, прежде всего, обнаруживаются изменения в костной системе. Задержка превращений хрящевой ткани в костную является значительным негативным фактором трансформации остеобластического костного мозга в красный, что, в свою очередь, приводит к нарушению морфофункционального становления лимфоидных органов, которая проявляется изменениями морфологического и биохимического составов крови.
2. Статус органов гемоимунопоэза у животных новорожденного периода
У новорожденных телят обнаруживается тесная взаимосвязь между пренатальным развитием организма и абсолютной и относительной массой органов кроветворения. В частности у новорожденных телят красной степной породы с живой массой, соответствующей породным показателям, абсолютная масса костной системы наибольшая, тогда как относительная – наименьшая, по сравнению с недоразвитыми животными. Телята, живая масса которых соответствует породным показателям, имеют абсолютную массу костной системы 6,20±0,25 кг, а относительную – 19,70±0,61%. У пренатально недоразвитых абсолютная масса костной системы снижается (5,25±0,21 кг), а относительная – возрастает (23,20±1,13%). Абсолютная масса костной системы зависит от пренатального остеогенеза. Апофизарное окостенение большого вертела бедренной кости у пренатально развитых суточных телят достигает 4,12±0,30%, тогда как у недоразвитых – 0,76±0,19%.
Зависимость функции гемоимунопоэза от пернатального остеогенеза оказывается определенным соотношением хрящевой и костной ткани в каждом костном органе. Количество хрящевой тканей в отдельных костных органах разное, что зависит от биомеханических нагрузок и пренатального развития животных. С увеличением живой массы у суточных животных, уменьшается количество хрящевой ткани в костных органах. Однако нарушение остеогенеза, в свою очередь, вызывает задержку хондрогенеза. При условии нарушения хондрогенеза не полностью реализуются генотипические возможности роста костных органов в длину, что влияет на количество красного костного мозга.
У новорожденных телят в костной системе содержится от 38 до 55% костного мозга или же 2,5-4,5 кг. Как правило преобладает красный костный мозг. Особенно его обилие отмечается в длинных трубчатых костях конечностей.
Желтый (жировой) костный мозг содержится только в костях метаподия телят. В рудиментарных костных органах недоразвитых телят выявляется прямая трансформация остеобластического костного мозга в желтый, минуя стадию красного.
Абсолютные и относительные показатели массы и объема костных органов зависят от степени пренатального развития новорожденного, а костный мозг у новорожденных телят красной степной породы составляет 3,0% от живой массы и 40-50% от массы костной системы.
Каждый костный орган образован губчатой и компактной костной тканью. Компактная костная ткань наиболее развита там, где костный орган максимально выдерживает нагрузку на сгиб. Губчатая костная ткань размещаются преимущественно в тех костных органах, в которых преобладают силы упругих деформаций. Компактная и губчатая костная ткань может быть грубоволокнистой (незрелой), ячейки которой заполнены остеобластическим (костеобразующим) и красным (гемопоэтическим) костным мозгом. Она характеризуется наличием сетчато-волокнистых коллагеновых волокон, остеоциты размещаются разнонаправленными, а межклеточное вещество мало минерализовано. Ретикуло-фиброзная костная ткань характерна для костных органов плодов и новорожденных животных, у которых также преобладает остеобластический и красный костный мозг. Следует отметить, что компактный слой костной ткани, диафизов длинных трубчатых костных органов образуется путем эндостального (прямого) остеогенеза. В камбиальном слое надкостницы сначала образуются отдельные островки (разнообразные формы, но у большинства продольно вытянутые) грубоволокнистой костной ткани. Ячейки между ними заполнены рыхлой соединительной тканью и кровеносными сосудами. Костеобразование отводится довольно интенсивно. К концу новорожденного периода костные балки значительно утолщаются, ячейки уменьшаются. Однако трансформации остеобластического костного мозга в красный не происходит, так как и образование хрящевой ткани.
Микроокружение красного костного мозга образуют ретикулярная ткань и сосуды микроциркуляторного русла, а также минерализованный компонент – груоволокнистая костная ткань. Увеличение хрящевой ткани в костных органах задерживает превращение остеобластического костного мозга в красный, что проявляется снижением показателей морфологического и биохимического составов крови, а клинически – анемией.
Для прижизненной оценки морфофункционального состояния органа универсального гемоимунопоэза необходимо проводить биопсию костной ткани в участке 4 сегмента грудины (в крайнем случае – в участке большого бугра плечевой кости). Из биоптата готовят гистологические срезы по общепринятой методике.
Снижение интенсивности превращения остеобластического костного мозга в красный отрицательно влияет на второй центральный орган гемоимунопоэза-тимус.
Зрелый тимус-это эпителиально-лимфоидный орган, доли которого состоят из отдельных зон, каждая из которых содержит лимфоидные клетки, чаще одного класса, и не лимфоидные, которые образуют необходимое микроокружение для созревания лимфоцитов. Процесс созревания лимфоцитов, особенно Т-клеток, сопровождается "включением" специальных генов. Известно, что на определенных этапах созревания Т-лимфоцитов (тимоцитов) происходит экспрессия антиген-специфических рецепторов, появление или отбор клеток, распознающих специфичные для тимуса продукты, которые необходимы для взаимодействия структур иммунной системы. На клетках появляется достаточное количество рецепторов, обусловливающих доставку лимфоцитов из кровообращения в лимфоидные структуры. В каждой дольке выделяют четыре зоны: субкапсулярную, кортикальную, собственно мозговую и зону периваскулярной соединительной рыхлой ткани. Разница между корковой и мозговой зонами в тимусе значительно более четкая по сравнению с другими лимфоидными органами.
Тимус новонардженных телят и ягнят имеет две (правую и левую) четные шейные, одну шейную нечетную (промежуточную) и нечетную грудную части. Парные шейные части начинаются от щитовидного хряща гортани, размещаются вдоль трахеи и перед входом в грудную полость сливаются в нечетную шейную (промежуточную), которая соединяется перешейком с грудной частью неправильной квадратной формы, содержащейся слева в переднем средостении.
Масса тимуса новорожденных животных разная. Она имеет внутривидовые и породные особенности. Абсолютная и относительная масса тимуса наиболее исследована у новорожденных телят красной степной породы. При этом его абсолютная масса у телят с высоким морфофункциональным статусом колеблется от 150 г до 200 г.
Следовательно, масса тимуса, его морфометрические показатели у суточных животных имеют значительные колебания. Прежде всего она зависит от пренатального развития новорожденных животных. У животных с меньшей или большей живой массой относительно породных показателей абсолютная масса тимуса меньше, как и относительная. У суточных телят с живой массой, соответствующей породным показателям относительная масса тимуса достигает 0,65%, тогда как с меньшей – 0,27%, а большей – 0,16%. В такой же закономерности изменяются линейные параметры (длина и ширина) тимуса.
Масса и линейные параметры у животных новорожденного периода, зависят от соотношения тканевых компонентов тимуса. У суточных телят красной степной породы в шейных долях тимуса содержится 88,10±1,50% лимфоидной ткани, а в грудной – 91,80±0,84%. При этом относительная площадь корковой зоны превалирует над мозговой и их соотношение достигает 1,82: 1-1,98: 1. через 10 суток жизни телят соотношение корковой и мозговой зон сужается (1,18:1-1,68:1). В мозговой зоне формируется значительное количество тимических Телец.
Количество межчастичной рыхлой соединительной ткани в тимусе зависит от пренатального развития новорожденных животных. Чем больше выявляется пренатальное недоразвитие новорожденного, особенно с нарушением остеогенеза, тем большее количество обнаруживается стромальных структур в тимусе. Кроме того, количество стромальных структур в данном органе зависит от акцидентальной (быстрой или временной) трансформации тимуса, которая наступает при длительных, тяжелых родах или при функциональных расстройствах органов дыхания (асфиксии), пищеварения (диспепсии). При этом происходит массовая миграция лимфоцитов в кровь, что проявляется резким уменьшением массы тимуса и ростом относительного количества стромальных структур. Акцидентальная трансформация тимуса-это проявление его высокой функциональной активности на негативное действие факторов как внутренней, так и внешней среды и является обратной реакцией. Пока еще не полностью исследована динамика структуры тканевых компонентов тимуса. У матуронатных животных в мозговой зоне обнаруживаются шаровидные образования, ретикулярная строма которых имеет аналогичность со строением корзинок лимфоидных узелков лимфатических узлов. Привлекает к себе внимание соответствующее образование сосудов микроциркуляторного русла в участках этих образований.
Селезенка является наиболее анатомически сформированным органом по сравнению с другими периферическими органами иммунной системы. Доказано, что основная ее функция заключается в нейтрализации в крови микробов, токсинов, погибших эритроцитов, пигментов и электроотрицательных коллоидов. Селезенка содержится на пути течения артериальной крови из магистральных сосудов (брюшной аорты, от которой ответвляется брюшная артерия) в воротную вену. Селезенку составляют строма (капсула и трабекулы, содержащие неполосованные миоциты), а также паренхима. Последняя представлена белой и красной пульпой. Белую пульпу формирует иммуноактивная часть селезенки, образованная лимфоидной тканью, способной выполнять функцию биологического надзора. Снаружи селезенка покрыта серозной оболочкой, под которой находится фиброзная капсула. От нее в толщу органа отходят трабекулы. Между ними расположена паренхима селезенки. В строме селезенки определяют ретикулярные клетки и волокна. В петлях ретикулярных клеток обнаруживаются плазмоциты, гистиоциты, лимфоциты и другие клетки, участвующие в иммунных реакциях.
Красная пульпа состоит из ретикулярной стромы и паренхимы (эритроцитов, лейкоцитов, лимфоцитов, макрофагов и других клеток). У телят селезенка длинная с закругленными концами серо-синего цвета, а ее относительная масса колеблется в пределах 0,3-1,2%.
В брюшной полости новорожденных телят селезенка расположена вдоль последних пар ребер. На ней определяют две поверхности: диафрагмальную и внутренность. На внутренностях находятся ворота селезенки, через которые в орган проникают сосуды и нервы. Снаружи селезенка покрыта брюшиной, связками которой она соединяется с желудком и диафрагмой. Такое анатомотопографическое расположение селезенки определяет одну из главных функций этого органа – иммунный надзор за течением крови.
Установлено, что у новорожденных телят площадь белой пульпы селезенки колеблется в пределах от 7,0 до 11,0%. Доказано, что клетки селезенки способны захватывать и обезвреживать микроорганизмы и другие посторонние факторы. Кроме того, в ней проходят процессы, которые несвязаны с иммунной защитой. В селезенке происходит гемолиз эритроцитов с освобождением железа, депонируется 16% крови, которая проходит сквозь красную пульпу.
Белая пульпа - это совокупность лимфоидной ткани, которая может быть диффузной (периартериальные связи) или узелковой, где происходят процессы контроля текущей крови, а также клеточная и субклеточная дифференциация, кооперативные взаимодействия т-и В-лимфоцитов, плазмоцитов и макрофагов в ячейках ретикулярной ткани. В лимфоидных узелках различают 4 не четко отграниченных зоны: периартериальную, центр размножения, мантийную и краевую (маргинальную).
Периартериальная зона занимает небольшой участок новколо центральной артерии и образована, главным образом, Т-лимфоцитами и интегрирующими клетками, которые адсорбируют антигены, попадающие с кровотоком и передающие Т-лимфоцитам информацию о состоянии микроокружения, стимулируя их бласттрансформацию и пролиферацию. В функциональном отношении периартериальная зона является аналогом паракортикальной тимусзависимой зоны лимфатических узлов.
Центр размножения или герминативный центр узелка образован ретикулярными клетками и пролиферирующими в-лимфобластами. Кроме этого, довольно часто здесь можно обнаружить скопления макрофагов с фагоцитированными лимфоцитами или их фрагментами. В таком случае центральная часть лимфоидного узелка выглядит светлой и является “реактивным центром”. На границе с мантийной зоной обнаруживаются плазмоциты. Функционально эта область идентична герминативным центрам лимфоидных узелков в лимфатических узлах.
Мантийная зона окружает периартериальную зону и центр размножения, образованная, главным образом, из плотно расположенных малых В-лимфоцитов и небольшого количества Т-лимфоцитов, а также содержащая плазмоциты и макрофаги.
Краевая зона представляет собой промежуточный участок между белой и красной пульпой шириной около 1000 мкм. Она содержит т-и в-лимфоциты и одиночные макрофаги, окружена краевыми или маргинальными, синусоидальными сосудами со щелевидными порами в стенке, через которые проникают лимфоциты и макрофаги.
Во вторичных лимфоидных узелках ретикулярные волокна образуют "корзину" значительно большего размера, образующую крупнозвездную сетку, несколько разрушенную в центре.
Характерно, что абсолютная масса селезенки суточных телят значительно меньше массы тимуса, а у 10-суточных телочек ее масса возрастает почти в два раза. Однако количество белой пульпы увеличивается незначительно. Среди лимфоидной ткани обнаруживаются первичные лимфоидные узелки (без центров размножения) и вторичные (с центром размножения) в незначительном количестве.
Лимфатические узлы расположены на пути течения лимфы по лимфатическим сосудам, протокам и стволам, выполняя функцию биологических фильтров. Лимфатические узлы в зависимости от их расположения разделяют на соматические (париетальные), внутренности (висцеральные) и смешанные, в которые поступает лимфа как от внутренних органов, так от париетальных структур. Форма лимфатических узлов у телят новорожденного периода разнообразна, но каждый из них имеет изогнутую и выпуклую поверхность. Через выпуклую поверхность лимфатические сосуды входят в узел, а через изогнутую (ворота органа) выходят.
В каждом лимфатическом узле, как самостоятельном органе, различают соединительнотканную основу, в которую входят капсула с воротным утолщением и трабекулы, отходящие от них, а также паренхиму, которая состоит из корковой и мозговой зон.
Капсула и трабекулы построены из плотных коллагеновых волокон, среди которых содержатся малочисленные фибробласты, фиброциты, неполосованные мышечные клетки и незначительное количество эластических волокон. Капсула и трабекулы образуют основу лимфатического узла. Наряду с этим есть еще более развитая строма из особо тонких ретикулярных волокон, которая заполняет пространства между трабекулами. Для каждой функциональной зоны лимфатического узла характерна особая сетка аргирофильных волокон. В мозговой зоне она мелковичковая, а в лимфоидных узелках – крупновичковая. Количество волокон колеблется в зависимости от участков лимфоидного узелка: их меньше в светлых центрах и больше на периферии. Таким образом, агрофильные волокна образуют" корзинки " характерного для лимфоидных узелков строения.
По морфологическим особенностям в паренхиме лимфатических узлов определяют корковую и мозговую зоны. Характерным для корковой зоны является наличие лимфоидных узелков, которые у телят могут иметь светлые центры. В висцеральных лимфатических узлах новорожденных телят лимфоидных узелков значительно больше, чем у соматических. Лимфоидные узелки со светлыми центрами обнаруживаются преимущественно у телят с высоким морфофункциональным статусом.
Мозговая зона образована мякотными тяжами, которые разветвляются и анастомозируют. В основе мозговых тяжей лежит ретикулярная ткань, в глазках которой содержатся в-лимфоциты, плазматические клетки и макрофаги.
Синусы, по которым течет лимфа, ограничены капсулой и трабекулами, с одной стороны, узелками и мозговыми тяжами – с другой. Мозговые синусы в области ворот сливаются в воротный синус, из которого выходят эфферентные лимфатические сосуды.
У суточных телят соматические лимфатические узлы имеют большую абсолютную массу и линейные промеры по сравнению с висцеральными. Относительная масса лимфатических узлов суточных телят колеблется в пределах сотых или тысячных долей процента. Характерно, что у 10-суточных телочек абсолютная и относительная масса лимфатических узлов изменяется незначительно, по сравнению с другими лимфоидными органами.
У суточных телят красной степной породы корковая зона несколько меньше по сравнению с мозговой. Однако количество диффузной лимфоидной ткани в корковой зоне превалирует, особенно в краниальном средостенном лимфатическом узле и узле подвздошной кишки (32,17-36,37%). Максимальное количество узелков без центров размножения, которые содержатся в корковой зоне, обнаруживается в краниальном средостенном лимфатическом узле (1,47-2,55%). Наличие лимфоидных узелков с центрами размножения указывают на способность лимфатических узлов давать определенный иммунный ответ на антигенные раздражители. Количество лимфоидной ткани в лимфатических узлах суточных телят составляет более 50%.
Следовательно, у суточных телят, лимфатические узлы сформированы и содержат не только диффузную лимфоидную ткань, но и лимфоидные узелки. Максимальное количество лимфоидных узелков в паренхиме больше в висцеральных лимфатических узлах по сравнению с соматическими. В корковой зоне лимфатических узлов обнаруживаются лимфоидные узелки со светлыми центрами, что свидетельствует об их реактивности. Наличие вторичных лимфоидных узелков у суточных телят возможно связано как с их матуронатностью, так и антигенной стимуляцией белками молозива. С возрастом животных количество лимфоидных узелков в лимфатических узлах значительно возрастает, что указывает на их активное участие в кооперации иммунной защиты организма.
Прижиттевую оценку морфофункционального состояния лиматических узлов рекомендуем проводить на базе биоптата поверхностного шейного лимфатического узла.
3. Определение организменного статуса новорожденных телят
На сегодня для создания здорового высокопродуктивного поголовья животных, необходимо, прежде всего, определять морфофункциональный статус организма новорожденных животных. Проводя многолетние исследования, мы разработали тесты для определения морфофункционального статуса организма у суточных телят по 20-балльной шкале (табл. 1).
Таблица 1
Тесты морфофункционального статуса организма новорожденных телят
|
№ |
Тест |
Абсолютная |
Бал |
Абсолютная |
Бал |
Абсолютная |
Бал |
|
1 |
Длина хвоста (см) |
0-2 |
6 |
3-5 |
3 |
6-8 |
1,5 |
|
2 |
Длина последнего ребра (см) |
До 5 |
4 |
5-7 |
2 |
Более 7 |
1 |
|
3 |
Время реализации позы стояния (мин.) |
До 30 |
2 |
До 60 |
1 |
Более 60 |
0,5 |
|
4 |
Количество резцовых зубов |
6-8 |
2 |
2-4 |
1 |
2 и меньше |
0,5 |
|
5 |
Время проявления рефлекса сосания (мин.) |
До 30 |
2 |
До 50 |
1 |
Более 50 |
0,5 |
|
6 |
Кожа, волосяной покров |
Эластичная; густой, блестящий, длинный |
1 |
Пониженная эластичность; густой, средней длины |
0,5 |
Собрана в складки, которые долго расплавляются; короткий, жидкий |
0 |
|
7 |
Количество эритроцитов (млн./ мкл) |
7 и более |
1 |
6-7 |
0,5 |
Меньше 6 |
0 |
|
8 |
Количество лейкоцитов (тыс. / мкл) |
8-9 |
1 |
6-8 |
0,5 |
Меньше 6 |
0 |
|
9 |
Живая масса (кг) |
30-35 |
1 |
25-29 |
0,5 |
Меньше 25 и более 35 |
0 |
|
Всего баллов |
20 |
10 |
4 |
Наибольшее количество баллов (10) мы уделили особенностям пренатального остеогенеза, который обеспечивает благополучие организма. Развитие костной системы, отдельных костных органов, в пренатальный период онтогенеза обеспечивает реализацию локомоторных актов у суточных животных, что необходимо для их выживания, а также становления органов гемоимунопоэза. При этом развитие костной системы можно определить визуально и промерами с использованием мерного оборудования (рис. 1). Наиболее информативными являются биометрические показатели рудиментарных костных органов (последняя пара ребер и хвоста), в которых максимально проявляются нарушения пренатального остеогенеза.
1. длина хвоста (6 баллов). При этом учитывается расстояние между кончиком хвоста (без волосяного покрова) и вершиной пяточного бугра (рис. 1). Заплесневелый сустав находится в состоянии максимальной экстензии, что соответствует обычному состоянию во время статики животного. Это расстояние у новорожденных телят колеблется от 0 до 7 см и более. Чем она меньше, тем больше жизнеспособность новорожденного теленка, что предопределяется наиболее полной реализацией генетических потенций роста и развития костной системы.
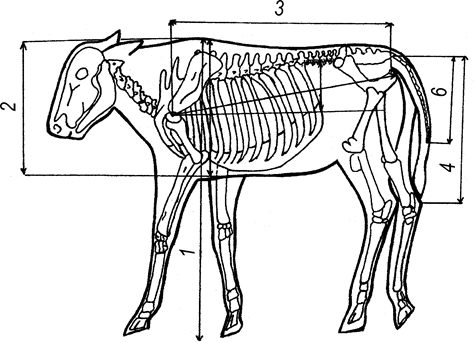
Рис. 1.
Основные промеры полов тела новорожденного теленка: 1-высота в холке; 2-глубина груди; 3-косая длина туловища; 4-Длина тазовой конечности от седалищного горба до вирховки пяточного горба; 5-длина последнего ребра; 6-Длина хвоста
При этом, рентгенографическим методом исследования определяется, что краниальные (головки) и каудальные (ямки) эпифизарные центры окостенения имеют первые 7 хвостовых позвонков, а диафизарные – 10. в костных органах (грудине, хвостовых позвонках, длинных трубчатых костях и т.п.) до 39,62±2,24–47,44±1,85% массы занимает красный костный мозг, что указывает на его высокую гемопоэтическую и иммуногенетическую активность.
Данный показатель отражает различную степень силы коррелятивных взаимосвязей у новорожденных телят между длиной хвоста и другими параметрами в участках туловища и конечностей, особенно, высотой в холке, глубиной грудной клетки и косой длиной туловища. Чем меньше расстояние между кончиком хвоста и вершиной пяточного бугра, тем сильнее взаимосвязи определяются между относительной массой костной системы, развитием сердца, легких, органов пищеварения и, особенно, иммунокомпетентных структур. Об условиях нарушения пренатального остеогенеза хвостовых позвонков расстояние от кончика хвоста до пяточного бугра возрастает, что можно определить визуально.
2. Длина последнего ребра (4 балла) у телят прогнозирует высокую жизнеспособность. При этом, учитывается расстояние между вентральным концом последнего ребра и фронтальной линией, которую условно проводят через плечевой сустав. У новорожденных телят это расстояние колеблется от 3 до 8 см. Чем больше длина ребра, тем меньше расстояние и, соответственно, больше развита костная система, выше жизнеспособность теленка. При этом, также учитывают коррелятивные связи между длиной последнего ребра и показателями полов тела в области туловища и конечностей. Чем выше коэффициент корреляции, тем выше жизнеспособность теленка. У пренатально недоразвитых телят последнее ребро почти не оказывается пальпацией и определить его длину практически невозможно. В таком случае балльная оценка приближается к нулю.
3. Время и характер реализации локомоторных актов (2 балла). Новорожденные телята, как представители матуронатного вида животных, сравнительно легко встают на конечности, реализуя позу стояния через 20-30 минут после рождения. Первые шаги шаткие и медленные. Сейчас тонус мышц и движения в пространстве становятся более уверенными. У недоразвитых телят после рождения реализация локомоторных актов задерживается. Движения их длительное время некоординированы.
4. Количество резцовых зубов (2 балла). При рождении у теленка прорезаются 6-8 резцов и 8 премоляров. Зубная формула их такова 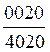 или
или ![]() .
.
Слизистая оболочка десен светло-розового цвета. У недоразвитых телят количество резцов меньше или они полностью отсутствуют. Подвижность резцовых зубов при пальпации также подтверждает недоразвитие костной системы. Красноватые слизистые оболочки ротовой полости и носогубного зеркальца также свидетельствуют о пренатальном недоразвитии особенно костной системы у телят.
5. состояние врожденных (безусловных) рефлексов (2 балла). У развитых, здоровых телят через 20-30 минут после рождения проявляется рефлекс сосания. С задержкой кормления во времени, телята могут сосать инородные предметы. Особенно такой феномен наблюдается при рождении теленка в поздний вечерний час при отсутствии обслуживающего персонала. В таком случае новорожденные потребляют подстилку или мочу, что приводит к нарушению функции органов пищеварения. Значительная задержка проявления во времени рефлекса сосания регистрируется у телят с невысоким морфофункциональным статусом организма.
Признаком пренатального недоразвития является запоздалое проявление рефлекса сосания (1-6 часов). Он дряблый, слабый или же вовсе отсутствует. Иногда такие слабые телята громко и часто ревут. Защитные реакции не выражены.
6. состояние кожного покрова (1 балл) взаимосвязано с морфофункциональным состоянием аппарата движения. У здорового новорожденного теленка длинные, густые и блестящие волосы, кожа умеренно влажная и эластичная. Образовавшаяся складка кожи на туловище быстро распрямляется. Снижение эластичности кожи особенно часто отмечается у животных с клиническим проявлением нарушения функции органов пищеварения. Телята с короткими и сухими волосами и кожей с пониженной эластичностью (образовавшаяся складка медленно выпрямляется), как правило, часто заболевают в первые часы и дни после рождения.
7. клеточный состав крови определяется в лабораторных условиях. У хорошо развитых телят количество эритроцитов больше, чем 7 млн/мкл (т / л), что составляет по тестовым показателям 1 балл. Количество гемоглобина колеблется в пределах 10,65-11,14 г/100 мл. Снижение его количества связано с нарушением гемоимунопоэтической функции костной системы, в частности костного мозга. От рождения и до первого кормления молозивом количество лейкоцитов более, чем 8 тыс/мкл (Г/л), достигая при тестовой оценке телят 1 балла. После своевременной кормежки молозивом их количество увеличивается до 12 тыс/мкл (Г/л), так же, как и гемоглобина. Следует отметить, что у новорожденных телят в первые дни после рождения обнаруживаются изменения и в соотношении лейкоцитов крови. К концу новорожденного периода возрастает количество базофилов, эозиофилов, лимфоцитов, моноцитов и сегментоядерных нейтрофилов на фоне уменьшения юных и палочкоядерных нейтрофилов. У недоразвитых телят красной степной породы количество эритроцитов в крови бывает меньше – 6,5 млн/мкл, а лейкоцитов до кормления молозивом – 7 тыс/мкл, а после – 8-9 тыс/мкл.
Установлено, что наряду с изменениями морфологического состава крови после рождения изменяется и ее белковый состав. На фоне роста в первые пять дней после рождения живой массы и, соответственно, объема плазмы крови происходит увеличение фетального гемоглобина и белка в сыворотке. Однако с возрастом у новорожденных телят уменьшается содержание альбуминов и, особенно, в первый день после рождения – более чем на 23%. Такая же закономерность проявляется в динамике α-и β-глобулинов. Характерно, что γ-глобулины в крови телят сразу после рождения не обнаруживаются, но уже в суточном возрасте их количество превышает аналогичную α - и β-глобулинов вместе (на 16,3%), уменьшаясь в дальнейшем, а показатели сыворотки крови телят уменьшаются с возрастом.
8. живая масса (1 балл) должна соответствовать средним показателям породы. В последнее время особенно у новорожденных телят, обнаруживается снижение живой массы на 3-10 кг, что является привычным явлением. Необходимо отметить, что низкая живая масса не всегда является показателем гипотрофии. В условиях ведения скотоводства, которое сложилось на сегодня, иногда рождаются также телята с живой массой, которая преобладает породные показатели. Увеличение живой массы обусловлено, прежде всего, накоплением в мышцах и фасциальных пространствах большого количества влаги, что также негативно влияет на жизнеспособность новорожденных животных.
При определении морфофункционального статуса новорожденных телят по рекомендуемым тестам их условно возможно поделить на три группы:
1 группа, организм которых имеет максимальное количество баллов (20) при оценке по поданным тестам. Это свидетельствует, что новорожденный теленок имеет высокий морфофункциональный статус и такую же жизнеспособность, что обусловлено полной реализацией генетических потенций пренатального роста и развития. Такие телята быстро адаптируются к условиям внешней среды, у них интенсивно проявляются условные рефлексы на время кормления, проживания, стадности и т.п. (рис. 2). В настоящее время на ферме в 500 голов коров рождается не более 7-15% таких телят. Для создания в будущем здорового молочного поголовья коров, с высокой степенью реализации генетических возможностей продуктивности, рекомендуется лишь таких телят использовать для восстановления родительского стада.
Во 2 группу выделяют телят с оценкой морфофункционального статуса организма в 10-19 баллов. У таких телят в период внутриутробного развития, особенно в последние недели плодового периода, происходит задержка не столько количественных изменений, сколько качественных. Полные изменения происходят в костных органах, и слизистая оболочка, особенно тонкой кишки, не может выделять достаточное количество кишечного сока для переваривания молозива. Как правило, на 2-3 сутки у них проявляется диспепсия. После лечения, с последующим выздоровлением, такие животные медленно растут, дают низкий прирост живой массы, даже при условии соответствующей кормежки и технологии выращивания (рис. 3).
Приплод, от таких животных, рождается еще с более низкой жизнеспособностью и, как правило, погибает во время рождения. В данном случае необходимо провести более широкие исследования морфофункционального статуса их организма по тестам с целью выявления дополнительных показателей нарушения их развития в пренатальном периоде онтогенеза. Одним из наиболее важных из них, как показывают наши исследования, является пренатальный остеогенез. Нередко проводят также клинические исследования родительского поголовья с целью исключения болезнетворных факторов, присутствующих в данном хозяйстве.

Рис. 2. Теленок и группы в Скотном дворе

Рис. 3. Теленок II группы в Скотном дворе
Выявив факторы нарушения пренатального роста и развития в организме телят, определяют комплексные мероприятия с целью профилактики. Главным условием в решении этой проблемы является введение в технологию выращивания и содержания животных на пастбищах с соответствующим количеством и качеством травостоя. При условии отсутствия пастбищ или в осенне-зимнее время года в технологии содержания животных использовать дозированное принудительное движение по терренкуру или специально оборудованной площадке или скотному двору. Для новорожденных телят необходимым условием при этом является назначение активного движения в сочетании с добавлением в молозиво (молоко) минеральных веществ, особенно солей кальция, с целью улучшения роста и развития костной системы. Недоразвитых телят выращивают с целью получения мясных продуктов. Использование их для воспроизводства поголовья является экономически бесполезным и приводит к созданию поголовья с нарушением функции полового аппарата, снижением продуктивности и рождением приплода еще с более низкой жизнеспособностью.
III группа новорожденных телят, морфофункциональный статус организма которых достигает лишь 4-9 баллов, являются практически нежизнеспособными они проявляют значительные признаки пренатального недоразвития. Первое кормление молозивом вызывает неадекватную реакцию вследствие внутриутробной сенсибилизации к белкам материнского организма. Таки телята погибают к концу 1-2 суток или болеют тяжелой формой диспепсии и погибают. Нарушения роста и развития (гипотрофия и гипопластия, особенно костных органов, обеспечивающих жизнеспособность) у таких телят возникают на более ранних этапах роста и развития по сравнению с телятами второй группы (рис 4). Провести коррекцию недоразвития организма таких телят, в условиях современных хозяйств, становится практически не решенной проблемой. Создание соответствующих условий, интенсивное использование иммуно -, биостимуляторов и антибиотиков в некоторой степени сохраняет и продолжает существование таких телят. Резкая смена условий среды при переводе их из родильного отделения в помещение для дальнейшего выращивания, обусловливает снова их заболевание с тяжелым течением, что завершается нарушением органов пищеварения и дыхания.

Рис. 4.
Теленок III группы в Скотном дворе
Поэтому выращивание недоразвитых телят с морфофункциональным статусом организма в 4-9 баллов экономически невыгодно.
Определение морфофункционального статуса новорожденных телят имеет большое значение не только при коррекции технологии кормления и выращивания, но и проведении вакцинации против инфекционных заболеваний. Вакцинация недоразвитых телят не обеспечивает устойчивого иммунитета даже в конце молочного периода.
Таким образом, многогранные особенности пренатального роста и развития новорожденных телят при всех взаимосвязях, и причинно-следственных взаимодействиях обеспечивают постоянство внутренней среды организма в ответ на опосредованное, через организм матери, действие различного рода факторов среды. Развитие плода в утробе матери, изолированного, в некоторой степени, плодными оболочками и жидкостью, определяют автономизацию его роста и развития. Только действие чрезвычайно дестабилизирующего фактора (или совокупности антропогенных факторов) на материнский организм, предопределяет изменение роста и развития плода. Особое значение при этом имеет плацентарный барьер, который, в первую очередь, реагирует на негативное влияние факторов на материнский организм, предопределяя нарушение роста и развития плода.
Следовательно, способы определения морфофункционального статуса новорожденных животных требует дальнейших разработок. Однако, на сегодня, возникает необходимость его определения с целью проведения корректировки технологии кормления и выращивания животных в новорожденный период онтогенеза.
4. Коррекция пренатального недоразвития телят новорожденного периода
Анализ научной и справочниковой литературы показывает, что наиболее разнообразные способы выращивания новорожденных млекопитающих, разработанные для телят. Главным фактором, побуждающим разработку различных способов выращивания телят является значительное снижение их жизнеспособности, а также уменьшение их количества на 100 коров, что приводит к значительным осложнениям по возрождению поголовья животных.
На сегодня наиболее широко распространена технология кормления новорожденных телят, морфофункциональный статус организма которых соответствует 20 баллам. После реализации первых локомоторных актов (20-30 минут после рождения) и проявления рефлекса сосания, телят необходимо кормить первым теплым молозивом из сосковой поилки. При этом выпаивают около 1,5-2,0 литров молозива. Необходимо также учитывать время рождения теленка до первого кормления. Почти в 80% случаев животноводы не проводят кормления телят через два часа после их рождения. Первое кормление новорожденного теленка проводят больше, чем через 6-8 часов, что характерно для больших ферм и при его рождении в поздний вечерний час. В таком случае необходимо уменьшить объем молозива почти на ½ при первом кормлении и где-то через 1-2 часа скормить остаток молозива (обязательно при температуре около +35...+36ºС). Это значительно уменьшит нагрузку значительным объемом молозива на сычуг не производя до вскрытия пищеводного желоба.
Для профилактики нарушений функции органов пищеварения предлагаем при кормлении телят из сосковой поилки разбавлять молозиво 50% раствором глюконата кальция (или физиологическим раствором) 1:1 или 1:0,5, что соответствует, в определенной степени, разбавлению его секретом слюных желез при кормлении под коровой. Это обуславливает более оптимальное его переваривание и обеспечивает организм кальцием, который необходим для запуска функций иммунных органов и структур, а также профилактирует, в некоторой степени, аллергические реакции, вызванные действием iG матери при неполноценном плацентарном барьере. В последующие кормления, с учетом морфофункционального статуса теленка, количество раствора уменьшают, а увеличивают количество молозива (молока). К концу вторых или третьих суток выпаивают, таким образом, только молозиво (молоко). Его суточная норма для развитого новорожденного теленка с высоким морфофункциональным статусом организма должна достигать 6-8 литров.
Потребность однократного выпаивания молозива новорожденному теленку также определяют с учетом живой массы, на один кг которой приходится 50 мл молозива. Таким образом, теленку живой массой 40 кг требуется два литра молозива на однократное кормление за сутки – 6.
Однако лучше первую-третью кормежку телят проводить подсосом под коровой. Во-первых, при самостоятельном сосании дойки вымени коровы у теленка выделяется большое количество секрета слюнных желез. Секрет не только разбавляет густое молозиво, но и, что очень важно, вспенивает его, предотвращая створаживание в сычуге. Во-вторых, вспененные секретом слюнных желез глотки молозива, не способны открыть пищеводный желоб и этим самым профилактируется образование казеина в рубце. И еще одна очень важная вещь-вместе с наполнением молозивом сычуга полностью удовлетворяется безусловный рефлекс сосания. При этом теленок высасывает такое количество молозива, которое отвечает его физиологическим потребностям.
Итак, кормление здоровых новорожденных телят надо проводить, как рекомендуют многие исследователи, не позднее двух часов после рождения. Это обусловлено тем, что, первое теплое (неподогретое) молозиво имеет высокую кислотность (50,0–60,0 Тº), обеспечивая создание оптимальной среды (рН 1-2) для функционирования и размножения в сычуге грамположительной молочнокислой микрофлоры, создавая условия для переваривания молозива. Кроме того, в нем находится максимальное количество иммунных глобулинов, которые являются "стартовыми" антигенами для самого быстрого включения в функцию собственных иммунокомпетентных структур. При этом, эпителиальные клетки (энтероциты) слизистой оболочки кишок, имеют свойство пропускать молозиво без ферментативного расщепления белков, особенно Ig. Количество белков в молозиве достигает 20%, наличие витамина А придает ему желтую окраску, а солей магния – свойство слабительного действия.
Состав молозива, динамику его составных частей, необходимо учитывать при кормлении животных его заменителями, что широко практикуется в животноводстве. Привлекает внимание использование для кормления телят заменителя цельного молока (молоко для телят по наименованию «Кальво стар экстра». При чем из 1 кг «Кальво стар экстра" можно получить 9-10 литров полноценного молока. Известно, что" Кальволак " производят на основе сухой подсырной и казеиновой сывороток. По биохимическому составу он соответствует такому молоку, а отдельное взятии витамины, микроэлементы преобладают его. Кроме того," Кальво стар экстра " незаменим при условии нарушения пренатального развития теленка или маститах коров.
Кормление телят с морфофункцональным состоянием в 20 баллов заменителями молозива рекомендуется проводить с четвертых суток после рождения. Сухой "Кальво Старт экстра" растворяют в чистой воде с 1:8 или 1:9 в эмальорированной посуде и скармливают телятам при t +38...+40°С. Количество такого раствора на одно кормление не должно превышать 1,5-2 литра. Со временем увеличивают как кратность кормления, так и количество использования "Кальво Старт экстра".
В случае рождения пренатально недоразвитых телят "Кальво Старт экстра" используют по 0,5 литра за одно кормление увеличивая кратность до 4-6 в сутки. При этом профилактируется образование в сычуге казеиновых сгустков, а энергетическая ценность и наличие витаминов и минеральных веществ обеспечивают их потребность организмом. Итак, пренатально недоразвитый теленок получает полноценный корм, который обладает легкой и высокой переваривающей способностью и профилактирует нарушение функции органов пищеварения с признаками диареи.
Важным фактором при выращивании здорового теленка является кратность кормления его в неонатальный период. Кормление новорожденных телят два раза в сутки (утром и вечером), практикуемое на скотоводческих фермах, не столько приводит к экономии физического труда, а сколько является фактором, который вызывает нарушение функции органов пищеварения, нанося ущерб. Рекомендуется, в первые 3-4 суток, телят кормить четыре раза, а до 30-суточного возраста – три раза. Только старших телят, при условиях полной адаптации к окружающей среде и наличии грубых кормов (доброкачественное сено, дерть и т.д.) – два раза в сутки. При долгих перерывах между кормлением у неонатальных телят может возникнуть не только нарушение функции органов пищеварения, но и искажение рефлекса сосания и активности движения. У телят старшего возраста чувство голода в долгих перерывах между кормлением проявляется меньше в связи с тем, что они уже поедают грубые корма.
Телят, особенно в неонатальный период, необходимо поить водой, не опасаясь, что это обусловит нарушение функции органов пищеварения. Возле воды в кормушке должны находиться минеральные добавки, содержащие соли кальция. Известно, что теленку в неонатальный и молочный периоды для развития костной системы вообще (также и гемопоэтического красного костного мозга в частности) необходимо 20,0–22,0 г кальция в сутки.
Грубые корма телятам вводят в рацион, начиная с 10-14 дня после рождения, что совпадает с началом функционирования рубца и способствует более интенсивному пищеварению в нем. По технологии выращивания новорожденных телят для обеспечения их оптимального роста и развития рекомендуется загонно-выгульное выращивание.
Для наиболее благоприятных условий для роста и развития теленка после первого кормления молозивом лучше его перевести в обору, крытую навесом, защищенную щитами от господствующих ветров, на сухую соломенную подстилку в большом количестве. Выращивание телят в скотных дворах лучше всего способствует реализации их генетической потребности в активном движении (рефлекс следования за матерью), и соответственно балансу расхода и достояния энергии.
При такой загонно-выгульной системе выращивания телят, наиболее оптимально также обеспечивается действие на их организм различных факторов внешней среды, имеющих значительные суточные колебания:
1. температуры воздуха.
2. относительной и абсолютной влаги воздуха.
3. скорости потоков воздуха.
4. активной вентиляции помещения.
5.солнечной энергии.
6. режима активного движения.
Ощущение холода или тепла является результатом совместного действия температуры и движения воздуха, его влаги, солнечной энергии и ее отражения от окружающих предметов. В утробе материнского организма рецепторы кожи теленка изолированы от действия любых факторов. В первые минуты после рождения запускается в работу кожный анализатор под действием многих факторов, в том числе и температурного. Новорожденный организм впервые использует свои механизмы для поддержания постоянной температуры, несмотря на ее колебания в окружающей среде. В условиях содержания телят в оборе, постоянное изменение температуры окружающей среды приводит к выработке устойчивых адаптивных механизмов, которые способны чувствительно реагировать на малейшие ее колебания.
В случае нахождения новорожденного организма в клетке в помещении с постоянной и сравнительно высокой температурой, что предлагают многие авторы, этот механизм не включается в работу. Происходит торможение замещения пренатальных структур на функциональные, характерные для животных неонатального и молочного периодов. При этом, происходит на первом этапе (до 10 суток) более интенсивный рост живой массы, а затем снижение на фоне хранения внутриутробных структур тканей. Кроме того, такие телята болеют при незначительном изменении температурного режима или скорости и влажности воздуха в неонатальный период. Особенно такие условия содержания телят негативно проявляются в молочный период заболеванием бронхопневмонией.
Известно, что в организме новорожденных телят тепло образуется в результате обменных процессов, которые обеспечивают поддержание жизненных функций вследствие биохимических реакций преобразования питательных веществ в энергию. Чем больше получает теленок молозива, тем больше в его организме вырабатывается тепла и энергии, что, в свою очередь, повышает двигательную активность и аппетит.
Вторым источником тепловой энергии является работа скелетных мышц, которая реализуется при локомоции телят. Именно она определяет потерю и приобретение энергии у неонатальных телят, как дорзомобильных животных. Включается общебиологический закон траты и достояния энергии – чем больше тратит организм энергии на локомоцию, тем интенсивнее растет ее достояние, тем лучше растет и развивается животное.
Часть энергии расходуется на образование тепла. Тепло из организма телят в новорожденный период расходуется благодаря испарению влаги с поверхности кожи и с выдыхаемым воздухом. Доказано, что общий расход тепла у телят путем испарения при окружающей температуре от +3° до +20°с равен 1,43 МДж/м2 за сутки. Площадь поверхности кожи новорожденного теленка определяется в квадратных метрах и равна 0,667 м2. К ней добавляют 30% влаги, выделяемой через органы дыхания.
Препятствием расхода тепла являются липиды и волосы кожи. Липиды кожи образуются, в основном, из подкожного жира, количество которого находится в прямой зависимости от пренатального развития теленка и условий выращивания. Если содержать теленка в помещениях при высокой температуре, то уменьшается количество выделения тепла, снижается аппетит, нарушаются механизмы терморегуляции. Нежелательна также очень низкая температура и, особенно, интенсивное движение воздуха. При этом, расходуется большое количество питательных веществ на образование тепла, а интенсивное движение воздуха обуславливает простудные заболевания. Выращивание телят в Скотном дворе с закрытой крышей при минимальном движении воздуха способствует образованию изоляционного слоя воздуха, который сохраняет тепло организма.
Следовательно, общая тепловая изоляция организма новорожденного теленка зависит от кожного покрова, кормления, активного движения и составляет 3,8 единиц, постепенно увеличиваясь до 5 единиц в 20-суточном возрасте. Показатель тепловой изоляции новорожденных телят, необходимый для определения критической температуры – это количество выделяемого тепла, которое равно его расходам. Критическая температура для новорожденных телят составляет+12°... + 15° С. При условии хорошего кормления, невысокой влажности воздуха и незначительном его движении, достаточном количестве сухой подстилки (соломы), критическая температура для неонатальных телят, снижается до +4°с. поэтому выращивание телят в скотных дворах с крытой крышей с использованием дозированного принудительного движения по терренкуру является наиболее оптимальным. На вторые сутки жизни теленку необходимо обеспечить активное движение. Для этого используют терренкур.
При выгульно-стойловом выращивании телят в первые сутки после рождения рекомендуется обеспечение естественной двигательной активности по скотному двору 20-30 минут до кормления. На четвертые сутки жизни телятам вводят дозированное принудительное движение по терренкуру, используя рефлекс следования за коровой-матерью (телятам несколько старшего возраста) со скоростью 35-40 м/мин. в течение 8-12 минут (300 г) (рис. 5). С 5-6 дня у телят уже вырабатывается рефлекс на дозированное принудительное движение. Расстояние увеличивают до 400-500 метров. После 10-суточного возраста телята хорошо пробегают до 1000 метров со скоростью 60-80 м/мин. Таким образом, выгульно-стойловая технология выращивания телят позволяет получать здоровых животных с использованием их, в будущем, для воспроизводства поголовья с высокими племенными качествами.
Если хозяйство не имеет скотного двора, то технология выращивания несколько меняется. Телятам дают возможность свободно двигаться по дорожкам помещения или на выгульных площадках. За содержание небольшого количества (1-20 голов) коров с использованием пастбищ, телят с 20-суточного возраста также рекомендуется содержать на пастбищах вместе с коровами.
Новорожденных телят, морфофункциональный статус которых составляет в 10-19 баллов (количество таких телят достигает 55-60% от всех новорожденных) даже при отсутствии клинических признаков какой-либо болезни, следует относить к группе риска, что подтверждается распространением среди них заболеваний в первые сутки жизни.
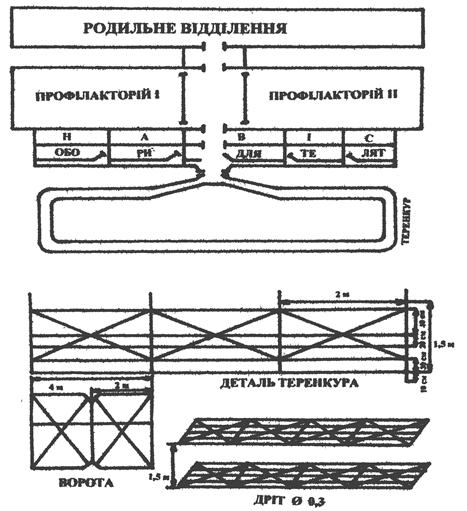
Рис. 5. Схема терренкура (100 м) для выращивания телят до 20-суточного возраста
Животные этой группы нуждаются в пристальном внимании непосредственно после рождения. Независимо от состояния здоровья, новорожденным телятам второй группы устанавливают диетический режим, не допуская выпаивания одного (чистого) молозива. При проявлении рефлекса сосания такому теленку выпаивают 0,5-1,0 литров 50% водного раствора глюконата кальция или отвары лекарственных растений (зверобой, ромашку, лекарственный тысячелистник, мяту, семена укропа, корни цикория и т.д.) за 30-60 мин. к кормлению молозивом. Затем выпаивают молозиво с глюкозо-солевым раствором глюконата кальция, увеличивая долю молозива в соотношении 5:1, 3:1, 2:1, 1:1 от первой выпойки до 5-6 после рождения.
При появлении клинических признаков нарушения функции органов пищеварения отменяют кормление молозивом, заменяя на 1-2 дня выпойкой глюкозо-солевого раствора глюконата кальция и травяных отваров в сочетании с парентеральной инъекцией раствора глюкозы, как антитоксического и энергетического средства. При этом следует учитывать, что наиболее эффективным является внутрикостное введение глюкозо-солевых растворов, (за исключением хлористого кальция), что позволяет инъецировать большие объемы растворов (до 1000-1500 мл) в относительно короткий срок. Способ внутрикостных введений очень прост при выполнении его в производственных условиях. Это не только уменьшает интоксикацию организма, но и обеспечивает его необходимым количеством воды.
В зависимости же от клинического состояния уже заболевших животных корректируют их содержание с лечением. Телят помещают в отдельную клетку с большим количеством соломы. Кроме того, окружающая температура должна быть не менее +20°...+22°С. В таких условиях они находятся 2-3 суток. Лишь на четвертые сутки в технологию содержания телят вводят дозированное принудительное движение.
Сначала такого теленка заставляют двигаться по клетке от 10 до 30 минут. Только после дозированного принудительного движения через 30 минут проводят кормление по определенной схеме диетотерапии. Через 3-5 суток, что зависит от клинического состояния, дозированное принудительное движение по терренкуру телятам применяют по предварительно указанной схеме. Телят по данной технологии содержат до месячного возраста. В соответствии с морфофункциональным статусом переводят на выгульное содержание. Телят с пренатальным недоразвитием в дальнейшем не рекомендуется использовать для воспроизводства поголовья.
Новорожденных телят, имеющих статус организма по тестовым показателям всего 4-9 балла, следует считать потенциально больными. Телята этой группы с первых суток после рождения характеризуются слабой двигательной активностью и проявлением основных рефлексов на фоне низкой жизнеспособности, что обуславливает их почти 100% заболеваемость с нарушением функции органов желудочно-кишечного тракта, аппарата дыхания и сердечной деятельности, при отсутствии возбудителей инфекционных болезней. Первое кормление молозивом таким телятам, независимо от их клинического состояния, нужно заменять глюкозо-солевым раствором глюконата кальция. В дальнейшем, до 3 суток после рождения, в солевой раствор добавляют незначительное (1/2-1/3) количество молозива. Такая диетотерапия предопределяет более быстрое становление пищеварительной функции. При первых признаках нарушения функции органов пищеварения телят третьей группы необходимо перевести на»голодную диету". При этом проводят систематическое лечение, применяя внутрикостное (внутришневенное) введение энергетических лекарственных веществ. В. И. Левченко не рекомендует, однако, заменять глюкозу при энтеральном введении сахарозой или крахмалом потому, что эти кормовые добавки способствуют углублению диспепсического синдрома.
Параллельно с выпаиванием глюкозо-солевого раствора больные телята нуждаются в растительных отварах и настоях (тысячелистник обыкновенный, конский щавель, зверобой обыкновенный, ромашка лекарственная), которые изготавливают по соответствующей схеме. В эмалированной емкости раздробленные части лекарственных растений заливают водой из расчета 1: 10.на водяной бане эту смесь выдерживают 30 минут и фильтруют через 2 слоя марли. Задают в дозе 10 мл/кг живой массы 2-3 раза в день в период голодания, в дальнейшем – за 30 минут до выпаивания молозива вместе с подтитрованными антибиотиками.
Кроме классических лекарственных средств оздоровления телят с организменым статусом 4-9 баллов, нужно использовать активную антисенсибилизирующую и стимулирующую терапию. При этом застовывают препараты заместительного действия, природные иммунокорректоры (витамины, препараты тимуса, костного мозга). Из препаратов заместительного действия используют также сыворотку крови коров, которую смешивают с глюкозо-солевым раствором в соотношении 1:1 или 1:3 и вводят интроперитониально в дозе 15-20 мл на кг живой массы. При наличии клинических признаков обезвоживания и интоксикации организма интраосально вводят 5% раствор глюкозы на физиологическом растворе 1000,0-1500,0 мл. Однако такие телята очень слабые и часто погибают даже при условии соответствующей терапии. В случае клинического выздоровления, у таких телят отмечается нарушение роста и развития. Животных этой группы содержат в отдельных станках или помещении с окружающей температурой +20°....+ 22°с, сухим воздухом и хорошим освещением.
Двигательную активность у недоразвитых телят с морфофункциональным статусом 4-9 балла назначают на 6 сутки после рождения. Дозированное движение таких телят также проводят сначала в станках. Теленок двигается 3-5 минут за полчаса до кормления. В последующие дни активное движение ежедневно увеличивают на 2-3 минуты. Только с 10-12 суток жизни таким телятам в технологию выращивания вводят дозированное принудительное движение по терренкуру или скотному двору по схеме, изложенной выше. Переболевшие телята значительно отстают в росте и развитии, а в будущем уменьшают реализацию генетических потенций и продуктивности на 30-35%.
Рекомендуемая литература
1. Анатомия домашних животных: учебник / С. К. Рудик, Ю. А. Павловский, Б. В. Криштофорова и др.; Под ред. С. К. Рудика. - К.: аграрное образование, 2001. - с.
2. клиническая диагностика внутренних болезней животных / В. И. Левченко, В. В. влезло, И. П. Кондрахин и др.; Под ред. В. И. Левченко. - Белая Церковь: БГАУ, 2004. - с. 456-472.
3. внутренние болезни животных / В. И. Левченко, И. П. Кондрахин, В. В. влезло и др.; Под ред. В. И. Левченко. - Белая Церковь: БГАУ, 2001. – ч. 2. - С. 313-333.
4. отчеты о выполнении научно-исследовательской работы №110/237-пр из НУБиП Украины по теме «исследовать особенности морфогенеза иммунокомпетентных структур и разработать критерии морфологической оценки их недоразвития во взаимосвязи с жизнеспособностью животных неонатального периода» за 2007-2009 года.