Костные органы
Костная система, как интегрирующая опорная конструкция организма, является одной из самых главных структур в иммунной кооперации защиты организма животных и человека в силу того, что ее неотъемлемой структурной частью является красный костный мозг.
Длительное время исследователи определяли, что центральным органом иммуногенеза является лишь красный костный мозг. Однако в последнее время доказано, что он является составной частью кости как органа [312]. А. Я. Фриденштейн [313] досказал, что неотъемлемой функцией костных органов является гемоиммунопоэтическая, которая выполняется ее структурным компонентом красным костным мозгом. Последний, как центральный орган иммуногенеза, является источником иммунокомпетентных стволовых клеток-предшественников, которые мигрируют в тимус (клоакальную сумку), а также в периферические органы иммуногенеза, где проходит их пролиферация и иммунологическое дозревание.
В соответствии с полифункциональностью костные органы имеют достаточно сложное строение. К структуре каждого костного органа входят губчатая и компактная костная ткань, костный мозг, ендост, периост, суставные и метафизарные хрящи.
В филогенезе кроветворную функцию костные органы, а именно костный мозг впервые начинает функционировать (и возникает) у животных, которые вышли из водной среды в наземную среду, – у амфибий. Выход позвоночных на сушу, а затем переход их к хождению и бегу, вызывает рост потребности организма у кислорода и предопределяет дополнение остеогенной функции костных органов кроветворной за счет красного костного мозга (П. А. Коржуев, 1966).
По данным ряда исследователей костный мозг, особенно красный, имеет достаточно сложное строение и имеет строму (ретикулярная ткань) и паренхиму (гемопоэтические клетки на разных стадиях дифференцирования), кровеносные сосуды, жировые клетки и нервы [347, 348]. Он имеет темно красный цвет, полужидкую консистенцию и заполняет костно-мозговые ячейки губчатой костной ткани, а также среднюю треть диафиза трубчатых костей.
Функционально в красном костном мозге различают миелоидную ткань (образует форменные элементы крови) и клетки лимфоидного ряда, совокупность которых можно рассматривать как его лимфоидные структуры. Необходимо отметить, что сложность методики исследования структуры костного мозга не дает возможности достаточно полно осветить динамику его строения, характер превращений с возрастом животных.
Однако, в настоящее время существуют доказательства его интенсивной трансформации. Так характерно, что у птиц после 15-суточного возраста (начало пневматизации костных органов) в костном мозге оказываются лимфоидные узелки. При этом формирование лимфоидных узелков происходит поэтапно. Сначала образуется скопление лимфоидной ткани среди красного костного мозга (рис. 10). Образование диффузной лимфоидной ткани в виде значительного скопления лимфоцитов больше всего оказывается в диафизе длинных трубчатых костей (особенно бедренной) конечностей.
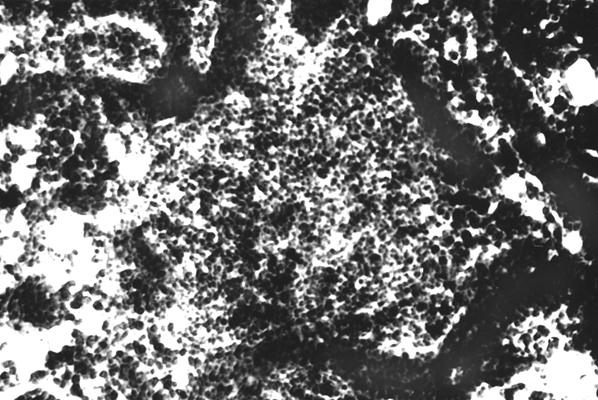
Рис. 10. Гистотопограмма диафиза бедренной кости курицы (180 суток) гематоксилин и эозин, МБИ-6, х80: 1 – диффузная лимфоидная ткань (Криштофорова Б. В., 1992).
На первом этапе образования лимфоидного узелка, вокруг диффузной лимфоидной ткани образуется сплетение из ретикулярных волокон, которые отмежевывают ее от красного костного мозга (рис. 10). С возрастом, в последующем образуются лимфоидные узелки, которые четко обмежевываются ретикулярными волокнами. Лимфоидные узелки имеют сферическую форму (рис. 11 и 12). Количество лимфоидных узелков растет. Они образуют скопления, в которых содержатся разной величины шаровидные лимфоидные узелки (рис. 14). По нашему мнению, наиболее реактивные лимфоидные узелки содержат розеткообразные центры (рис. 15). Не исключена возможность наличия подобных образований в красном костном мозге млекопитающих. Сложность выявления лимфоидных узелков, на наш взгляд, связанная с массивностью костных органов и отсутствия методики выработки гистотопограмм, которые полностью отображали бы их структуру. Наши исследования показывают, что как скопление лимфоидной ткани, так и лимфоидные узелки, оказываются в диафизах длинных трубчатых костных органов щенков собак.
Исследования показывают, что относительная масса костного мозга к живой массе составляет 4,5-4,7%, а к массе скелету – 40-45%. На первом этапе образуется остеобластический (костеобразующий) костный мозг, который трансформируется в красный (гемоиммунопоэтический). С возрастом животных красный костный мозг замещается желтым (жировым). Красный костный мозг у половозрелых животных остается в грудине, ребрах и позвонках в пограничной зоне с хрящевой тканью.
А. Я. Фриденштейн, А. С. Лалыкина [311] досказывают наличие тесной функциональной связи между костным мозгом и костной тканью минерализованным компонентом микроокружения. Получив чистую культуру клеток паренхимы костного мозга и пересадив ее под капсулу почки опытного животного, авторы установили, что из нее образуется сферическая кость, в которой происходит гемопоез.
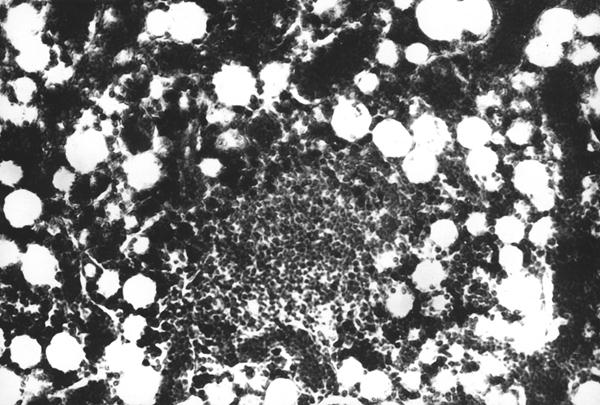
Рис. 11. Гистотопограмма диафиза бедренной кости курицы (180 суток) гематоксилин и эозин, МБИ-6, х80: 1 – первый этап формирования лимфоидного узелка (Криштофорова Б. В., 1992).
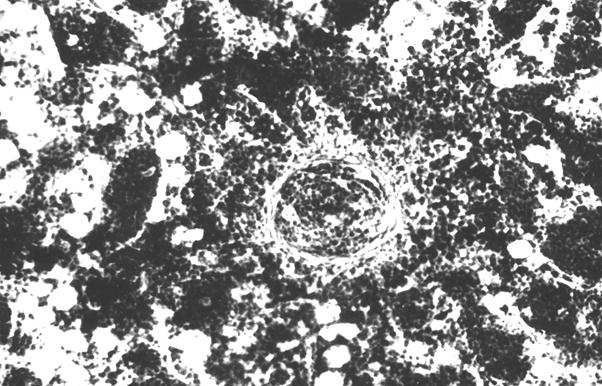
Рис. 12. Гистотопограмма диафиза бедренной кости курицы (240 суток) гематоксилин и эозин, МБИ-6, х80: 1 – лимфоидный узелок (Криштофорова Б. В., 1992).
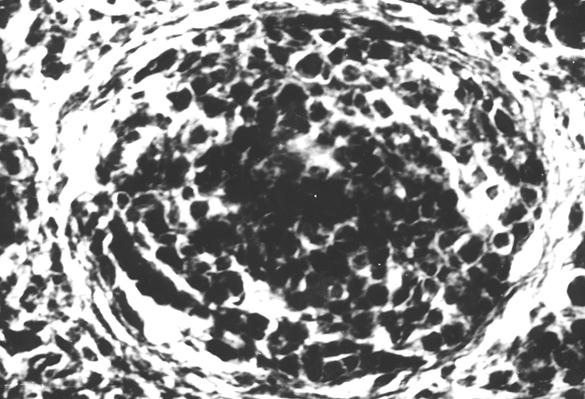
Рис. 13. Гистотопограмма диафиза бедренной кости курицы (240 суток) гематоксилин и эозин, МБИ-6, х400: 1 – лимфоидный узелок, окруженный ретикулярными волокнами (Криштофорова Б. В., 1992).
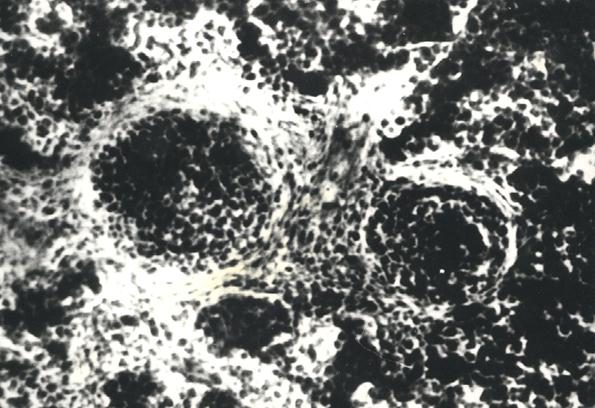
Рис. 14. Гистотопограмма диафиза бедренной кости курицы (240 суток) гематоксилин и эозин, МБИ-6, х80: 1 – лимфоидные узелки (Криштофорова Б. В., 1992)
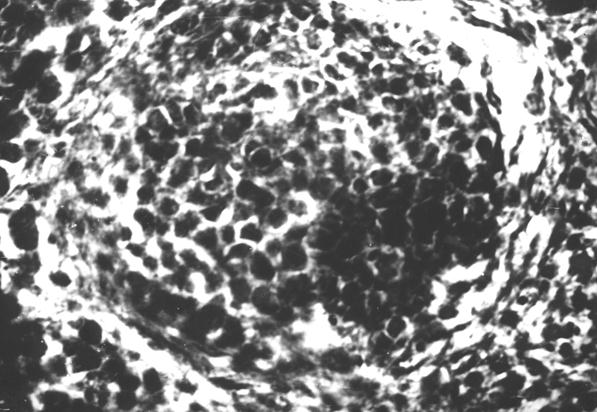
Рис. 15. Гистотопограмма диафиза бедренной кости курицы (240 суток) гематоксилин и эозин, МБИ-6, х400: 1 – лимфоидный узелок с 2 – розетковидными центрами (Криштофорова Б. В., 1992).
Костный мозг, как неотъемлемая часть кости как органа, становится главным звеном, что связывают организм в единственное целое с окружающей средой. У млекопитающих костный мозг определяется как один из центральных органов иммуногенеза, который поставляет ствольные полипатентные клетки для всех предшественников миело - и лимфоцитопоэза.
Строма красного костного мозга образована ретикулярной тканью и элементами микроокружения, что характерно для других органов гемоиммунопоэза. Паренхиму красного костного мозга образуют ствольные клетки и клетки миелоидного и лимфоидного рядов, которые находятся на разных стадиях развития [348]. На протяжении суток в организме животного образуется 250-300 млрд. эритроцитов, что подтверждает большое функциональное значение красного костного мозга для организма. Следует отметить, что образование клеточных структур в костном мозге проходит в соответствии с потребностями организма и реализацией генетических потенций развития. Количественная потребность его реализуется на уровне дифференцирования клеток, которые потеряли полипотентность, но еще способных к размножению.
Установлено, что у новорожденных животных красный костный мозг пронизан большим количеством капилляров, которые составляют до 50% от его массы и способствуют проникновению в общее кровообращение зрелых клеток [77, 78].
Кроветворение у новорожденных животных, как и у всех млекопитающих, происходит за пределами сосудов. Только зрелые и способные к миграции клетки-предшественники проходят через стенку синусоидов (расширенных посткапилляров) и венул в общее кровообращение. Доказано, что усиление кровотока в костном мозге способствует эритроцитопоэзу, а снижение – гранулоцитопоэза. В связи с этим необходимо обеспечивать животных двигательной активностью, особенно матуронатних, с первых суток жизни.
Формирование и размещение красного костного мозга проходит поэтапно. На первом этапе происходит разрушение трабекул первичной губчатой ткани, на фоне которой формируются одиночные скопления кроветворных клеток (рис. 16). С образованием трабекул вторичной губчатой ткани одиночные скопления кроветворных клеток увеличивается и сливается в коловидные образования небольшого размера (рис. 17). Разрушение первичной губчатой и костной ткани предопределяет размещение красного костного мозга отдельными скоплениями среди клеток остеобластичного. В последующем с трансформацией ретикулофиброзных костных балок в пластинчатую костную ткань происходит значительный рост отдельных коловидных образований красного костного мозга, которые почти заполняют костномозговые воротнички вторичной губчатой костной ткани в грудных позвонках и грудине (рис. 18, 19). Однако следует отметить, что в одной костномозговой ячейке оказываются как центры красного костного мозга, так и одиночные скопления кроветворных клеток, что больше всего характерно для костных органов новорожденных животных (рис. 20). У 5-суточных поросят подобные коловидные скопления красного костного мозга оказываются уже и в грудине (рис. 21). С возрастом животных, в 10-суточных, скопление красного костного мозга имеет более четкую границу (рис. 22). Возможно, такие образования скоплений красного костного мозга связаны со значительным дифференцированием полипотентных клеток, которое приводит к интенсивной динамике морфологического состава крови.
Следует отметить, что рядом со скоплениями красного костного мозга, уже у 20-суточных поросят происходит интенсивная его трансформация в желтый параллельно слизистым разрушением костных трабекул (рис. 23).

Рис. 16. Гистотопограмма позвоночной части последнего ребра поросенка (сутки). Гематоксилин и эозин, МБИ-6, х80: 1 – разрушенная трабекула первичной губчатой костной ткани; 2 – остеобластический костный мозг; 3 – хрящевая ткань (В. Г. Соколов, 2003).
В красном костном мозге между кровеносными сосудами, миелоидные и лимфоидные образования размещаются в виде шнуров. Гемопоэтические клетки содержатся между кровеносными сосудами, с одной стороны, и эндоостом с второй, гранулоциты – в центре образования костно-мозгового тяжа и, лишь на стадии образования мета - и миелоцитов, приближаются к стенке синусоидных гемокапилляров. Тромбоциты образуются, из отделенных полосок цитоплазмы мегакариоцитов, уже в просве синусоидов. Лимфоциты и моноциты размещаются вокруг ветвей артерий, особенно артериол, которые обеспечивают начало выполнения функции обмена веществ.

Рис. 17. Гистотопограмма 8-го грудного позвонка поросенка (сутки). Гематоксилин и эозин, МБИ-6, х80: 1 – скопление красного костного мозга (В. Г. Соколов, 2003).
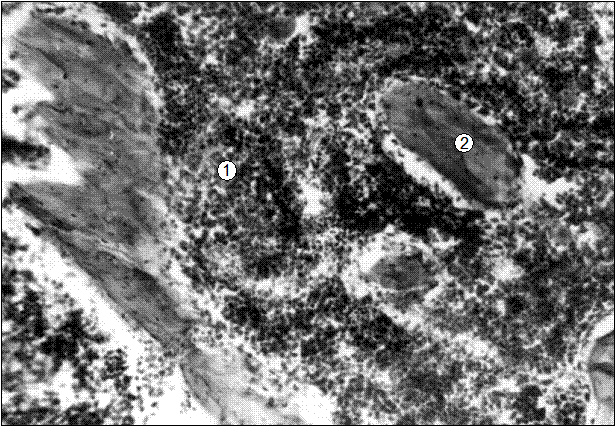
Рис. 18. Гистотопограмма 8-го грудного позвонка поросенка (сутки). Гематоксилин и эозин, МБИ-6, х80: 1 – скопление красного костного мозга; 2 – трабекула вторичной губчатой костной ткани (В. Г. Соколов, 2003).

Рис. 19. Гистотопограмма 4-го сегмента грудины поросенка (сутки). Гематоксилин и эозин, МБИ-6, х80: 1 – красный костный мозг; 2 – трабекула вторичной губчатой костной ткани (В. Г. Соколов, 2003).

Рис. 20. Гистотопограмма средней части диафиза 3-пястной кости поросенка (сутки). Гематоксилин и эозин, МБИ-6, х80: 1 – красный костный мозг; 2 – адипоцит (В. Г. Соколов, 2003).

Рис. 21. Гистотопограмма 4-го сегмента грудины поросенка (5 суток). Гематоксилин и эозин, МБИ-6, х80: 1 – красный костный мозг; 2 – трабекула вторичной губчатой костной ткани (В. Г. Соколов, 2003).

Рис. 22. Гистотопограмма 4-го сегмента грудины поросенка (10 суток). Гематоксилин и эозин, МБИ-6, х80: 1 – красный костный мозг; 2 – трабекула вторичной губчатой костной ткани (В. Г. Соколов, 2003).
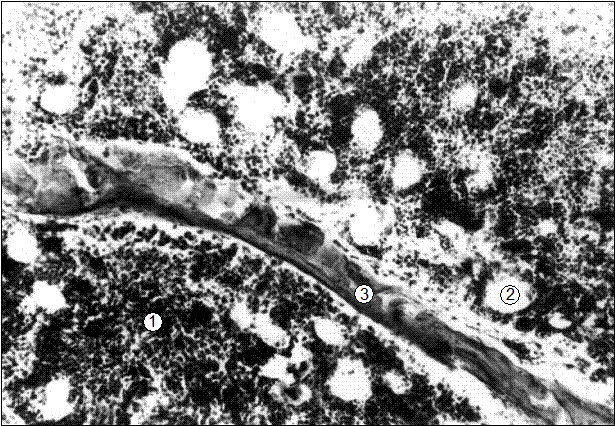
Рис. 23. Гистотопограмма 8-го грудного позвонка поросенка (20 суток). Гематоксилин и эозин, МБИ-6, х80: 1 – красный костный мозг; 2 – адипоцит; 3 – трабекула вторичной губчатой костной ткани (В. Г. Соколов, 2003).
Концентрация ствольных кроветворных клеток увеличивается от центра костномозгового участка в направлении эпифизов костных органов. Ствольные клетки преимущественно содержатся рядом с ендостом, что является доказательством выполнения костной тканью функции микроокружения и поддержки популяции активного гемопоеза [49].
Лимфоциты, которые образуются в костном мозге составляют 5-20% от всех клеточных популяций, поступают через синусоидные капилляры в кровь, а затем в тимус и другие лимфоидные органы.
У новорожденных животных оказывается тесная взаимосвязь между пренатальным развитием организма и абсолютной и относительной массой органов кроветворения (табл. 4). Ж. Г. Стегней (Грабчак) [76] доказывает, что у новорожденных телят красной степной породы с живой массой соответственно породным показателям, абсолютная масса костной системы наибольшая, тогда как относительная – наименьшая, сравнительно с недоразвитыми животными (табл. 5). Так, телята, живая масса которых отвечает породным показателям, имеют абсолютную массу костной системы 6,20±0,25 кг, а относительную – 19,70±0,61%. У пренатальных недоразвитых – абсолютная масса костной системы снижается (5,25±0,21кг), а относительная – растет (23,20±1,13%). Как показывает автор, абсолютная масса костной системы зависит от пренатального остеогенеза. Апофизарные окостенения большого вертлюга бедренной кости у пренатальных развитых суточных телят достигает 4,12±0,30%, тогда как в недоразвитых – 0,76±0,19%. Ряд авторов [16, 65, 68, 133, 134, 149] показывает, что пренатальный онтогенез влияет на развитие всего организма.
Таблица 4.
Взаимосвязь между живой массой и некоторыми кроветворными органами у новорожденных телят (Ж. Г. Стегней (Грабчак), 1998)
|
Показатели |
Живая |
Масса скелета |
Апофизарные окостенения % |
Масса тимуса |
Масса селезенки |
|||
|
Абсолютная, кг |
Относительная % |
Абсолютная, |
Относительная % |
Абсолютная, г |
Относительная % |
|||
|
М±м |
23,14±1,88 |
5,25±0,21 |
23,20±1,13 |
0,76±0,19 |
62,89±20,19 |
0,26±0,07 |
38,80±6,08 |
0,16±0,02 |
|
Minash max |
15-30 |
4,35-6,00 |
20,00-29,00 |
0,36-1,61 |
21,07-171,00 |
0,10-0,66 |
24,05-70,86 |
0,10-0,27 |
|
V,% |
19,92 |
9,71 |
11,94 |
61,84 |
78,66 |
65,38 |
38,40 |
31,25 |
|
М±м |
25,14±1,32 |
5,52±0,15 |
21,72±0,52 |
2,80±0,13 |
79,43±21,79 |
0,30±0,08 |
46,45±7,26 |
0,18±0,02 |
|
Minash max |
22-32 |
5,12-6,22 |
19,44-23,27 |
2,54-3,23 |
34,33-192,57 |
0,13-0,71 |
26,24-85,72 |
0,12-0,32 |
|
V,% |
12,85 |
6,70 |
5,84 |
11,43 |
66,72 |
66,67 |
38,30 |
27,78 |
|
М±м |
31,83±2,24 |
6,20±0,25 |
19,70±0,61 |
4,12±0,30 |
86,33±25,11 |
0,26±0,06 |
61,76±7,62 |
0,19±0,02 |
|
Minash max |
26-39 |
5,56-6,99 |
17,92-21,39 |
3,48-5,54 |
35,52-205,10 |
0,14-0,53 |
34,08-80,60 |
0,13-0,25 |
|
V,% |
15,77 |
9,03 |
6,95 |
16,26 |
65,16 |
50,00 |
27,64 |
26,32 |
Таблица 5.
Количество хрящевой ткани в некоторых костях телят
с разной живой массой (Б. В. Криштофорова, В. В. Смоляк, 2000) %
|
Группы животных по морфофункциональному статусу организма |
И |
ІІ |
ІІІ |
ІV |
V |
VІ |
|
Живая масса |
19,62±0,81 |
26,00±0,58 |
27,00±1,87 |
32,66±0,33 |
37,33±2,60 |
38,00±0,58 |
|
Бедренная кость |
21,01±0,79 |
19,43±1,46 |
25,49±2,54 |
24,07±1,56 |
20,79±1,27 |
17,94±0,67 |
|
Грудина |
31,08±0,82 |
36,28±2,66 |
39,49±2,36 |
38,61±2,44 |
29,79±2,50 |
32,84±1,57 |
|
Последнее ребро |
17,03±1,19 |
16,25±1,48 |
16,42±1,46 |
15,47±1,05 |
14,07±2,71 |
11,15±1,71 |
|
Хвостовые позвонки (1-7) |
47,07±1,54 |
49,00±0,53 |
50,72±1,43 |
46,07±2,81 |
40,19±1,30 |
45,18±1,69 |
Зависимость функции гемоиммунопоэза от пренатального остеогенезу проявляется определенным соотношением хрящевой и костной ткани в каждом костном органе, на что указывают рентгенологические исследования, проведенные Б. В. Криштофоровой и В. В. Смоляком [153]. Результаты исследований приведены в таблице 4. Количество хрящевой ткани в отдельных костных органах разное, что зависит от величины биомеханических нагрузок и пренатального развития животных.
Мы отмечаем, что с увеличением живой массы у суточных животных, уменьшается количество хрящевой ткани в костных органах. Однако следует помнить, что нарушение остеогенеза в свою очередь вызывает задержку хондрогенеза. При условии нарушения хондрогенеза не полностью реализуются генотипичные возможности роста костных органов в длину, которая влияет на количество красного костного мозга.
У новорожденных телят в костной системе содержится от 38 до 55% костного мозга или же 2,5-4,5 кг. Как правило, преобладает красный костный мозг. Особенно его большое количество отмечается в длинных трубчатых костях конечностей.
Желтый (жировой) костный мозг содержится лишь в костях метаподия, что характерно для новорожденных жеребят и телят, тогда как у поросят и щенков он отсутствует.
Большое количество остеобластического костного мозга и хрящевой ткани оказывается в костных органах недоразвитых и имматуронатных животных. Кроветворную функцию последних в некоторой степени выполняет и печень (В. В. Лемещенко, 2000). Кроме того, в рудиментарных костных органах недоразвитых телят оказывается прямая трансформация остеобластичного костного мозга в желтый, минуя стадию красного [68].
В настоящее время не существует единственного взгляда на степень развития костной системы в целом и костного мозга в частности. В. В. Смолье [281] доказывает, что абсолютные и относительные показатели массы и объема костных органов зависят от степени пренатального развития новорожденного. П. М. Гаврилин [68], исследуя динамику этих показателей у новорожденных телят красной степной породы с живой массой соответственно породным показателям установил, что костный мозг составляет 30,0% от живой массы и 40-50% от массы костной системы.
Мы уже отмечали, что сложная структура костных органов характерна для всех млекопитающих. Каждый костный орган образован губчатой и компактной костной тканью. Компактная костная ткань наиболее развитая там, где костный орган максимально выдерживает нагрузку на изгиб. Губчатая костная ткань размещается преимущественно в тех костных органах, в которых преобладают силы упругих деформаций. Компактная и губчатая костная ткань может быть ретикулофиброзной (незрелой), ячейки которой заполнены остеобластическим (костеобразующим) и красным (гемопоэтичным) костным мозгом. Она характеризуется наличием сетчатоволокнистых коллагеновых волокон, остеоцити розмещаються разнонаправлено, а межклеточное вещество, слабо минерализовано. Ретикуло-фиброзная костная ткань характерна для костных органов плодов и новорожденных животных, у которых также преобладает остеобластический и красный костный мозг (рис. 24, 25). Следует отметить, что компактный слой костной ткани, диафизов длинных трубчатых костных органов образуется путем ендостального (прямого) остеогенеза. В камбиальном слое надкостницы сначала образуются отдельные островки (разнообразных форм, но в большинстве продольно вытянутые) грубоволокнистой костной ткани. Ячейки между ними заполнены рыхлой соединительной тканью и кровеносными сосудами (рис. 26). Костеобразование происходит достаточно интенсивно. До конца новорожденного периода костные балки значительно утолщаются, ячейки уменьшаются (рис. 27). Однако трансформации остеобластичного костного мозга в красный не происходит, так как и образование хрящевой ткани.
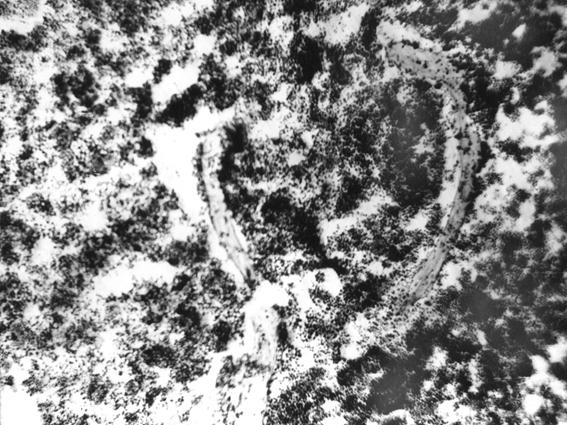
Рис. 24. Гистотопограмма грудины телочки (сутки) с высоким морфофункциональным статусом организма (В. В. Смоляк, 2000): 1 – красный костный мозг.
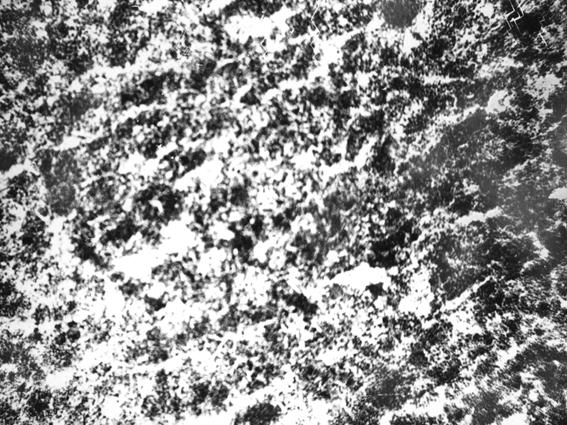
Рис. 25. Гистотопограмма грудины телочки (сутки) с пренатальным недоразвитием (В. В. Смоляк, 2000): 1 – красный костный мозг.
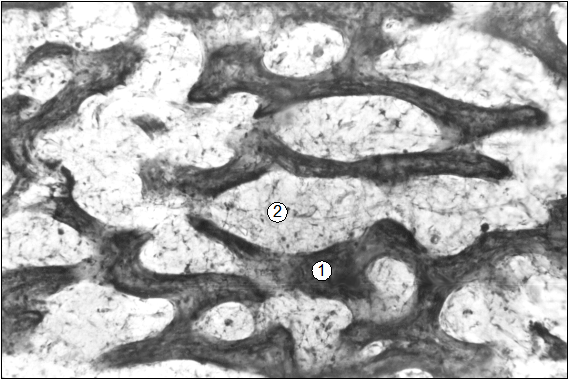
Рис. 26. Гистотопограмма средней части диафиза бедренной кости поросенка (сутки). Гематоксилин и эозин. МБИ-6, х80: 1 – трабекула компактной КТ; 2 – рыхлая волокнистая соединительная ткань.
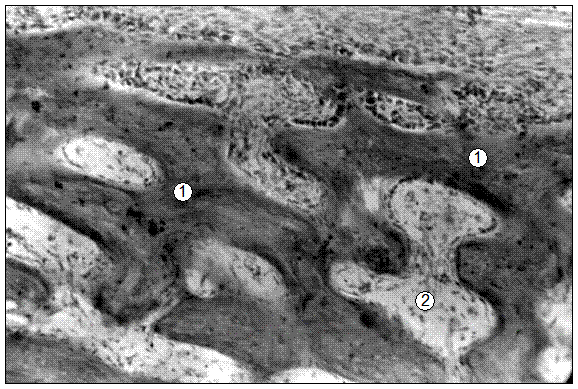
Рис. 27. Гистотопограмма средней части диафиза бедренной кости поросенка (10 суток). Гематоксилин и эозин. МБИ-6, х80: 1 – трабекула компактной КТ; 2 – рыхлая волокнистая соединительная ткань.
В средней трети диафизов позвонков все ячейки губчатой костной ткани заполнены красным костным мозгом (рис. 28). Однако уже у 10-суточных поросят среди красного костного мозга оказываются одиночные жировые клетки (рис. 29). Происходит перестройка самих костных трабекул – они приобретают пластинчатую структуру.
Зрелая (пластинчатая) костная ткань появляется с возрастом животных и характеризуется наличием остеонов, в которых содержатся коллагеновые волокна, на которых откладывается кристаллический кальций и значительно больше минеральных веществ. Такая костная ткань имеет более выраженные биомеханические свойства, а в ее костно-мозговых ячейках и средней части диафиза трубчастых костей содержится желтый костный мозг, который принимает непосредственное участие в биомеханической функции костного органа.
Хрящевая ткань (гиалиновая), образует суставные и метафизарные хрящи, толщина которых зависит от интенсивности пренатального остеогенеза. Исследования А. В. Олияр и В. Г. Соколова [234, 283-287] также подтверждают значительную зависимость количества красного костного мозга от интенсивности пренатального остеогенеза и положения костного органа в костной системе поросят.
У суточных поросят количество красного костного мозга в костных органах осевого скелету (позвонках), гораздо меньше сравнительно со скелетом конечностей (бедренная кость). Однако превращение красного костного мозга в желтый, с возрастом животных, быстрее проходит в костных органах скелету конечностей (рис. 28, 29).
В грудине суточных поросят наиболее информативно проявляется пренатальное становление красного костного мозга.
По данным Ю. Г. Смирновой [275] у новорожденных поросят костный мозг составляет 4,36% от живой массы и 26,0-32,0% от массы скелету, что предопределено интенсивностью пренатального развития. При уменьшении количества костного мозга кроветворная функция компенсируется печенкой. У новорожденных поросят костный мозг содержится преимущественно в осевом скелете (табл. 6, 7). Недостаточность функции красного костного мозга способствует возникновению анемии [110].
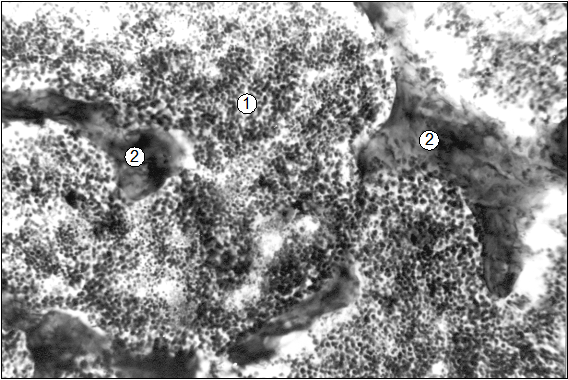
Рис. 28. Гистотопограмма диафиза 9 грудного позвонка поросенка (сутки). Гематоксилин и эозин. МБИ-6, х80: 1 – ККМ; 2 – трабекули вторичной губчатой КТ.
Рис. 29. Гистотопограмма диафиза 9 грудного позвонка поросенка (10 суток). Гематоксилин и эозин. МБИ-6, х80: 1 – ККМ; 2 – адипоцит; 3 – трабекулы вторичной губчатой КТ.
Таблица 6.
Относительная масса костного мозга новорожденных поросят к массе скелета (Ю. Г. Смирнова, 1968)
|
Живая масса, г |
Костный |
Относительная масса костного мозга % |
|||
|
Черепа |
Грудной конечности |
Тазовой конечности |
|||
|
1200,0 |
56,50 |
26,0 |
39,0 |
27,6 |
20,0 |
|
1170,0 |
45,44 |
18,2 |
27,2 |
31,8 |
28,5 |
|
1100,0 |
44,20 |
32,5 |
28,7 |
20,6 |
22,7 |
|
950,0 |
43,96 |
27,0 |
14,8 |
31,2 |
28,6 |
|
930,0 |
37,88 |
23,3 |
14,7 |
24,1 |
25,0 |
|
В среднем 1070 |
43,60 |
25,4 |
24,9 |
27,16 |
25,0 |
Таблица 7.
Динамика ВП костного мозга и его микроокружения в теле 9 грудного позвонка поросят (А. В. Олияр, 2002)
|
Возраст, |
ОКМ* |
ЧКМ* |
ЖКМ* |
Хрящевая |
Костная |
|
1 |
9,74±0,23 |
31,96±0,38 |
– |
30,75±0,25 |
23,78±0,21 |
|
5 |
8,19±0,11 |
35,13±0,15 |
– |
29,42±0,17 |
24,86±0,08 |
|
10 |
7,41±0,12 |
37,38±0,17 |
0,35±0,15 |
27,44±0,15 |
23,99±0,15 |
|
15 |
6,64±0,14 |
37,55±0,19 |
0,74±0.22 |
25,19±0,18 |
24,58±0,24 |
|
20 |
7,64±0,27 |
35,16±0,71 |
0,95±0,16 |
27,72±0,89 |
24,65±0,48 |
|
Бедренной кости |
|||||
|
1 |
8,79±0,33 |
34,68±0,59 |
0,18±0,03 |
26,61±0,44 |
22,21±0,51 |
|
5 |
6,00±0,51 |
39,57±0,16 |
1,13±0,07 |
24,74±0,62 |
25,57±0,39 |
|
10 |
5,68±0,32 |
37,89±0,49 |
2,87±0,27 |
23,36±0,45 |
25,85±0,48 |
|
15 |
4,71±0,25 |
38,14±0,34 |
3,39±0,23 |
23,28±0,23 |
27,50±0,44 |
|
20 |
6,16±0,52 |
36,18±0,65 |
3,06±0,27 |
25,73±0,62 |
26,32±0,51 |
Примечание: * ОКМ, ЧКМ, ЖКМ – остеобластический, красный и желтый костный мозг.
Следовательно, в организме новорожденных животных количество красного костного мозга составляет 3-5% живой массы или возле 50% абсолютной массы от костной системы. Микроокружение красного костного мозга образуют ретикулярная ткань и сосуды микроциркуляторного русла, а также минерализованный компонент – ретикулофиброзная костная ткань. Увеличение хрящевой ткани в костных органах задерживает превращение остеобластичного костного мозга в красный, что проявляется снижением показателей морфологического и биохимического состава крови, а клинически – анемией [110]. Кроме того, снижение интенсивности превращения остеобластичного костного мозга в красный негативно влияет на второй центральный орган гемоиммунопоэза – тимус.