Костная система
Костная система, как интегрирующая опорная конструкция организма, является одной из главных структур в иммунной кооперации защиты организма. Ее неотъемлемой составной частью является красный костный мозг, выполняющий функцию центрального органа гемоиммунопоэза. В костных органах в процессе фило- и онтогенеза костный мозг проходит 3 стадии развития. Первая стадия - остеобластический или костеобразующий костный мозг, который появляется только у костных рыб, что приводит к образованию фиброзной (грубоволокнистой) костной ткани. При наличии грубоволокнистой костной ткани происходит трансформация остеобластического костного мозга в красный, который выполняет функцию гемоиммунопоэза. Возможно, структура кости как органа и биомеханические функции, которые она выполняет в условиях земной гравитации, являются теми факторами, которые не только определяют интенсивность ее гемоиммунопоэтической функции, но в значительной мере обеспечивают наиболее оптимальные условия для ее выполнения (П. А. Коржуев, 1961, 1971).
Красный костный мозг имеет вид массы густой консистенции красного цвета, содержится в глазках вторичной губчатой костной ткани и костномозговых участках диафизов трубчатых костей конечностей. У суточных телят и поросят в этой области среди красного костного мозга встречаются единичные жировые клетки или их скопления, что является доказательством превращения его в желтый (рис. 3). Стромальной структурой костного мозга является ретикулярная ткань, а также остеогенные, жировые, адвентициальные клетки и макрофаги, которые наряду с сосудами микроциркуляторного русла образуют его микроокружение. Характерно, что для функционирования красного костного мозга необходим минерализованный компонент - ретикулофиброзная костная ткань. На клеточном уровне красный костный мозг - это совокупность полипотентных стволовых клеток, которые находятся на разных стадиях созревания: дифероны эритроидного, гранулоцитарного, мегакариоцитарного рядов, а также предшественники Т- и В-лимфоцитов. Созревшие клетки эритроидного и лимфоидного рядов через синусоидные капилляры, имеющие значительный поперечник (до 300 мкм и более), попадают в общий кровоток, обеспечивая морфологический состав крови. Отсюда следует вывод - состав крови, который исследуют при различных заболеваниях животных и человека, в полной мере зависит от функционального состояния красного костного мозга, каждого костного органа, всей костной системы в целом.
Характерно, что количество костного мозга, особенно красного, зависит от площади очагов окостенения, которые образуются еще в пренатальном периоде онтогенеза. Относительная площадь очагов окостенения, в свою очередь, зависит от интенсивности пренатального остеогенеза, что отражается на развитии всего организма. Так, площадь очагов окостенения всегда больше у телят, по сравнению с поросятами и, особенно, щенками. Колебания площади очагов окостенения наблюдается у животных одного и того же вида. У суточных поросят с признаками пренатального недоразвития в бедренной кости на рентгенограммах обнаруживается задержка пренатального остеогенеза, доказательством чего является отсутствие апофизарного очага окостенения большого вертела, а также проксимального эпифиза этого костного органа (рис. 4), тогда как у нормально развитых наблюдаются все основные и дополнительные очаги окостенения (рис. 5).
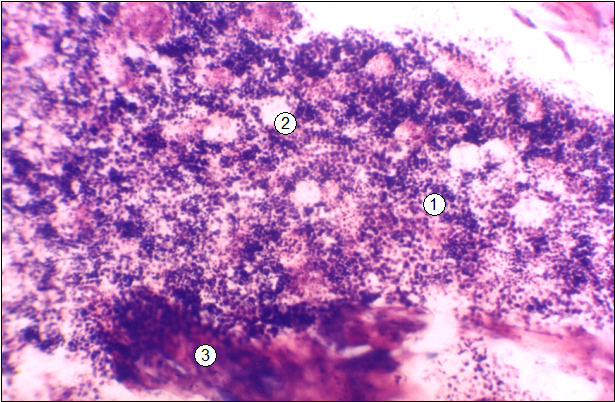
Рис. 3. Гистотопограмма среднего участка диафиза бедренной кости поросенка (сутки). Гематоксилин и эозин, МБИ-6, х80: 1 - красный костный мозг; 2 - адипоцит; 3 - трабекула вторичной губчатой костной ткани.
Рис. 4. Рентгенограмма бедренной кости недоразвитого поросенка (сутки): 1 - очаг окостенения диафиза; 2 - проксимальный эпифизарный хрящ; 3 - апофизарный (большого вертела) хрящ.
Рис. 5. Рентгенограмма бедренной кости поросенка (сутки): 1 - очаг окостенения диафиза; 2 - очаг окостенения проксимального эпифиза; 3 - очаг окостенения апофиза (большого вертела).
На тканевом уровне определяется прямая коррелятивная зависимость между площадью очагов окостенения костных органов и количеством красного костного мозга. У пренатально развитых животных (телят, поросят, щенков) при увеличении площади очагов окостенения увеличивается количество красного костного мозга и костной ткани и уменьшается количество остеобластического костного мозга и хрящевой ткани. У пренатально недоразвитых животных в органах универсального гемоиммунопоэза возрастает количество хрящевой ткани и остеобластического костного мозга.
В костных органах животных соотношение количества тканевых компонентов интенсивно изменяется в первые 20 суток жизни (новорожденный период) (рис. 6). Компактная костная ткань имеет характерное сетчатое строение, особенно в позвонках (рис. 7). Среди красного костного мозга в телах позвонков обнаруживаются единичные жировые клетки (рис. 8), количество которых возрастает в трубчатых костях конечностей, особенно в их диафизе (рис. 9). Рентгенографические исследования показывают значительное возрастание площади очагов окостенения как в осевом скелете (позвонки), так и скелете конечностей (рис. 10, 11).
Следует помнить, что в организме новорожденных (суточных) животных количество красного костного мозга составляет 3-5% от живой массы или около 50% от массы костной системы. Это связано с образованием значительного количества клеток для обеспечения ими крови. У млекопитающих наибольшее его количество содержится в костных органах осевого скелета - до 50,50%, а в скелете конечностей: тазовых - 38,70%, грудных - 10,80%. У птиц, наоборот: наибольшее количество костного мозга имеется в скелете конечностей - 62,50%, в осевом же - 8,50%. Функция желтого костного мозга на сегодня не определена. Предполагают, что он усиливает деформационные возможности костных органов. Его количество возрастает с появлением пластинчатой (зрелой) костной ткани, упругие возможности которой значительно ниже, по сравнению с грубоволокнистой (незрелой).
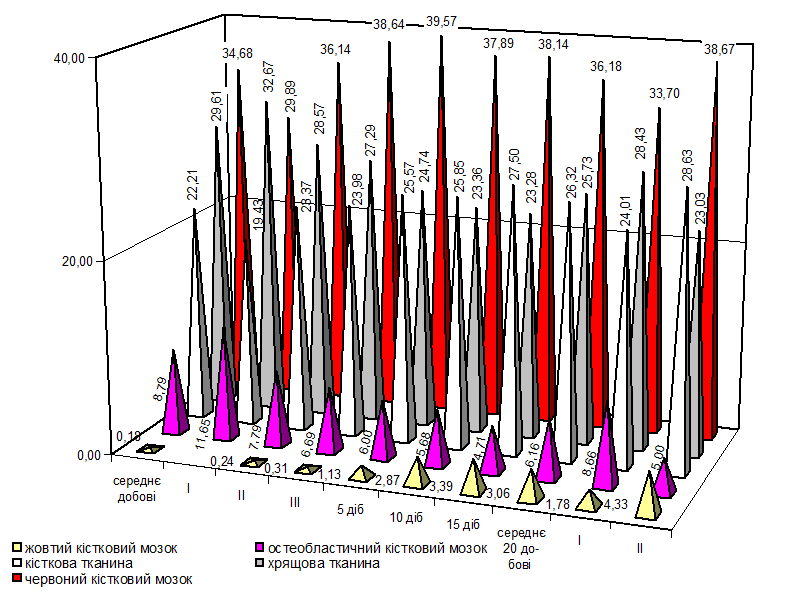
Рис. 6. Гистограмма динамики тканевых компонентов бедренной кости поросят.
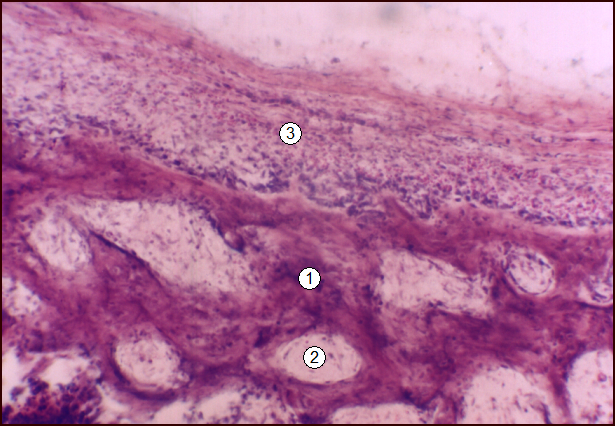
Рис. 7.Гистотопограмма 9-го грудного позвонка поросенка (10 суток). Гематоксилин и эозин, МБИ-6, х80: 1 - трабекула компактной костной ткани; 2 - рыхлая волокнистая соединительная ткань; 3 - периост.
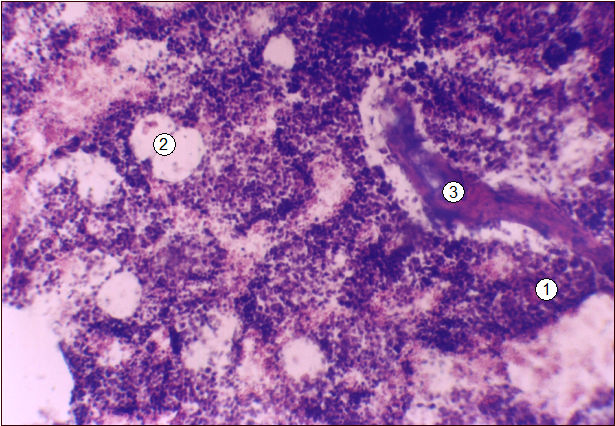
Рис. 8. Гистотопограмма диафиза 9-го грудного позвонка поросенка (10 суток). Гематоксилин и эозин, МБИ-6, х80: 1 - красный костный мозг; 2 - адипоцит; 3 - трабекула вторичной губчатой костной ткани.
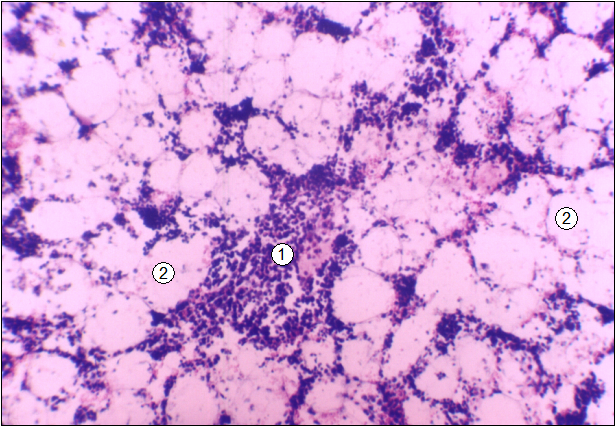
Рис. 9.Гистотопограмма среднего участка диафиза бедренной кости поросенка (20 суток).Гематоксилин и эозин, МБИ-6, х80:1 - красный костный мозг; 2 - желтый костный мозг.
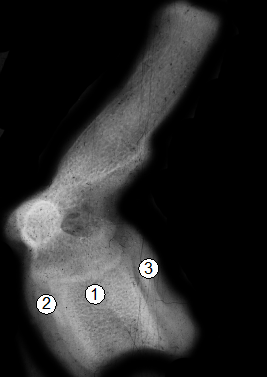
Рис. 10. Рентгенограмма 9-го грудного позвонка поросенка (20 суток): 1 - очаг окостенения диафиза; 2 - очаг окостенения краниального эпифиза (головки); 3 - очаг окостенения каудального эпифиза (ямки).
Рис. 11. Рентгенограмма бедренной кости поросенка (20 суток): 1 - очаг окостенения диафиза; 2 - очаг окостенения проксимального эпифиза; 3 - очаг окостенения апофиза большого вертела).