Механизм сокращения мышечных тканей
В молекулярном механизме сокращения мышечных тканей выделяют 2 процесса. Один из них кальций-зависимый, другой - АТФ-зависимый.
В поперечнополосатой и гладкой мышцах кальций-зависимый процесс осуществляется по-разному, а АТФ-зависимый – одинаково.
Кальций-зависимый прОЦесс сокращения в Исчерченной мышце
Сокращение мышечного волокна (или кардиомиоцита) происходит только в том случае, когда на актине открываются участки для связывания миозина, в результате чего миозин соединяется с актином. При этом этапность инициации сокращения происходит в следующей последовательности: сократительный стимул (нервный импульс) проходит по сарколемме и поступает на мембраны Т-трубочек, что стимулирует образование из липидов мембран Т-трубочек инозитол-фосфатов, которые взаимодействуют с рецепторами на мембранах АЭС, что инициирует открытие кальциевых каналов в её мембранах. Выход кальция из АЭС в цитозоль (в покое концентрация кальция в цитозоле 10-7-10-8 ммоль/л, при сокращении - 10-5) и его диффузия к миофибриллам завершается образованием комплексного соединения кальция с тропонином С, в результате чего на актине открываются места для связывания миозина, который соединяется с актином и происходит сокращение.
Кальций-зависимый процесс сокращения в гладкой мышце
Сокращение миоцита происходит вследствие фосфорилирования легкой цепи миозина, только в этом случае головка миозина может связывать и расщеплять АТФ и взаимодействовать с актином. Поступление сократительного стимула (нервный импульс, гормон) инициирует открытие кальциевых каналов в цитомембране миоцита, АЭС и митохондриях. Поступающий через эти каналы кальций соединеняется с кальмодулином. Образующийся комплекс кальций-кальмодулин активирует киназу легких цепей миозина, которая фосфорилирует легкие цепи его головок, в результате чего они приобретают способность связывать и расщеплять АТФ и соединяться с актином.
АТФ-зависимый процесс сокращения
Головка миозина присоединяет молекулу АТФ и расщепляет её до АДФ и фосфата, вследствие чего головка миозина присоединяется к актину. Одновременно с этим от головки миозина отсоединяются АДФ и фосфат. Именно в этот момент головка миозина делает гребковое движение и молекула миозина продвигается вдоль молекулы актина (другими словами – молекула миозина тянет на себя актин) (см. рис. 13). Вслед за этим головка миозина присоединяет новую молекулу АТФ и только после этого отсоединяется от актина и приобретает первоначальное положение.
Таким образом, без АТФ мышца не может ни сократиться, ни расслабиться.
В процессе сокращения мышечных тканей важную роль выполняет гладкая эндоплазматическая сеть (АЭС). В структурных единицах мышечных тканей (особенно в поперечнополосатой) очень хорошо развита АЭС, являющаяся хранилищем ионов кальция. В её мембране имеются кальциевые каналы, по которым кальций выходит из полости АЭС и входит обратно. Выход ионов кальция из плости сети в цитозоль осуществляется пассивно, так как их концентрация в цитозоле намного ниже, чем в полости АЭС, а поступление обратно в полость представляет собой активный транспорт с затратой энергии АТФ.
Сокращение поперечнополосатых мышц
В процессе сокращения поперечнополосатых мышц длина актиновых и миозиновых филаментов не изменяется, а происходит их смещение относительно друг друга: миозиновые нити вдвигаются в пространства между актиновыми, а актиновые - между миозиновыми; в результате этого: ширина I-диска и H-полоски А-диска уменьшается, в то же время ширина диска А не изменяется, а длина саркомера укорачивается (см. рис. 12).
В структурных единицах исчерченных мышечных тканей АЭС оплетает каждую миофибриллу и близко подходит к Т-трубочкам. Сигналом для выхода кальция из канальцев АЭС являются специальные регуляторные вещества – липидные медиаторы – инозитол-3-фосфат и инозитол-4-фосфат, которые синтезируются в мембранах Т-трубочек только в момент прохождения по ним сократительного импульса. Так как канальцы сети близко подходят к Т-трубочкам, инозитол-фосфаты быстро достигают АЭС и взаимодействуют со специфическими рецепторами для инозитол-фосфатов, находящимися на её мембранах. Это приводит к открытию кальциевых каналов в мембранах АЭС и быстрый выход ионов кальция из полости канальцев в цитозоль, что и инициируют сокращение.
Сокращение гладких мышц
В отличие от поперечнополосатых мышц, в которых миофибриллы существуют постоянно, в гладких мышцах они образуются только в момент сокращения, которое происходит вследствие поступления сигнала от нервных клеток. Под воздействием медиатора в плазмолемме миоцитов образуются кавеолы, в которые путём эндоцитоза поступают ионы кальция, вызывающие полимеризацию миозина и его взаимодействие с актиновыми филаментами. Актиновые филаменты одним своим концом с помощью сшивающих белков прикрепляются к специальным областям внутренней поверхности плазмолеммы, а другим - к миозину. Миозиновые филаменты прикрепляются к специальным местам в цитозоле клетки (нексусы).
|
|
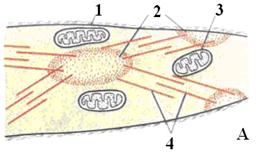
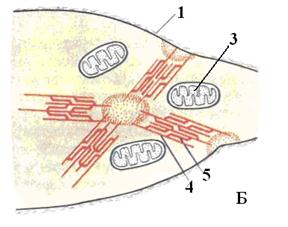
Смещение актиновых филаментов относительно миозиновых приводит к укорочению клетки. После прекращения поступления сигнала кальций покидает кавеолы, миозин деполимеризуется, миофибриллы распадаются и клетка расслабляется (рис. 14).
Рис. 14. Схема строения миоцита гладкой мышечной ткани в расслабленном состоянии (А) и при сокращении (Б). 1 – цитолемма: 2 – плотные тельца; 3 – митохондрии; 4 – актиновые филаменты; 5 - миозиновые филаменты.