Общая физиология микроорганизмов
Химический состав микроорганизмов
Химический состав микроорганизмов рассматривается на двух уровнях: Элементарном и молекулярном.
Элементарный уровень подразумевает пропорцию входящих в состав бактериальной клетки элементов (табл.1).
Таблица 1. Приблизительный элементарный состав бактериальных клеток (Р. Стейниер, Э. Эдельберг, Дж. Ингрем,1979)
|
Элементы |
Содержание, % от сухого вещества |
Элементы |
Содержание, % от сухого вещества |
|
Углерод Кислород Азот Водород Фосфор Сера Калий |
50 20 14 8 3 1 1 |
Натрий Кальций Магний Хлор Железо Все остальные элементы |
1 0,5 0,5 0,5 0,2 ~0,3 |
Всего микробная клетка может содержать около 90 различных элементов. Среди них выделяют две категории: Макроэлементы – те, которые содержатся в количестве более, чем 0,01%. К ним относятся, в первую очередь, Органогены: углерод, водород, кислород и азот. Суммарное их содержание в клетке около 90% и они составляют основу органического вещества. Далее следуют сера, фосфор, калий, натрий, кальций, магний и железо.
Вторая категория - микроэлементы – Входят в состав клетки в количествах менее 0,01%. К ним относятся йод, бор, молибден, цинк, медь, ванадий, серебро и др.
Молекулярный уровень Учитывает состав молекул. Все вещества микробной клетки делят на 2 группы : сухие вещества и воду - Н2О. Содержание воды в микробной клетке – 80-90%. Сухие вещества подразделяются на зольные (минеральные соли) – 2-5% и органические – белки, жиры, углеводы, нуклеиновые кислоты (табл.2).
Таблица 2. Содержание органических веществ в клетках бактерий ( по Stouthamer, 1973).
|
Название вещества |
Количество, % от сухих веществ |
|
Белки Углеводы Липиды РНК ДНК |
52 17 9 16 3 |
Механизмы поступления веществ в микробную клетку
Клеточная стенка имеет крупноячеистое строение и барьером при поступлении молекул не является. Существует лишь один основной барьер - плазмалемма, которая обладает избирательной проницаемостью (одни вещества пропускает, а другие - нет).
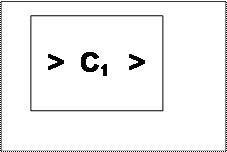
Между содержимым клетки и внешней средой всегда существует разница концентраций молекул и ионов. Этот перепад называется Электрохимическим градиентом И Является Двигателем перемещения веществ через мембрану. Если концентрация молекул или ионов за пределами клетки больше, чем в клетке (С2 > С1), такой раствор называется гипертоническим. Если же наоборот (С1 > С3), внешний раствор называется гипотоническим. Перемещение частиц из области большей концентрации в область меньшей называется движением по градиенту. Перемещение в противоположном направлении – движением против градиента концентраций и зарядов (рис. 19).
КЛЕТКА
 |
С2 С3
Рис.19. Схема перемещения молекул и ионов через клеточную мембрану: С1 – концентрация молекул в клетке;
С2 – концентрация гипертонического раствора; С3 – концентрация гипотонического раствора.
Перемещение частиц: по электрохимическому градиенту;
Против электрохимичекого градиента.
Поступление веществ в клетку зависит от многих факторов: соотношения их концентраций внутри клетки и за ее пределами, от размера молекул, энергетического состояния клетки и др. Выделяют два основных пути транспорта веществ через клеточную мембрану: пассивный и активный:
Пассивный транспорт Проходит по электрохимическому градиенту (из области большей концентрации молекул и зарядов в область меньшей концентрации) без затрат энергии извне (за счет энергии самого градиента).
Виды пассивного транспорта:
1. Простая диффузия – это движение через поры в мембране за счет осмотических сил, которое ограничивается только размерами пор;
2. Облегчённая диффузия - осуществляется при помощи белков – переносчиков (пермеаз), в том числе, по турникетному механизму.
Турникетный белок захватывает молекулу субстрата и за счёт энергии электрохимического градиента поворачивается на 180º. Здесь комплекс распадается и белок - переносчик возвращается в исходное положение, т. к. химическое сродство к наружной стороне мембраны у него выше, чем к внутренней (рис. 20).
ПЛАЗМАЛЕММА
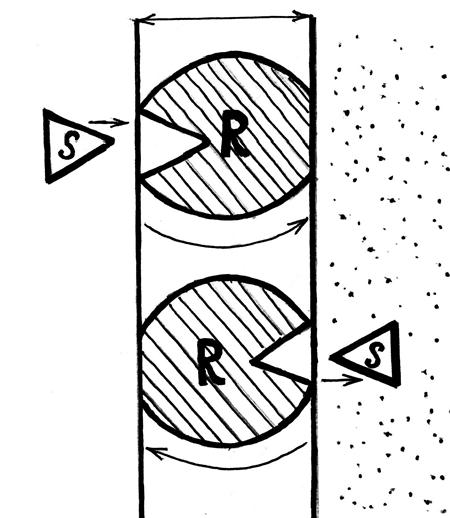 |
Рис. 20. Схема пассивного поступления веществ в клетку через плазмалемму по турникетному механизму. S - субстрат (молекула питательного вещества); R – мембранный белок-переносчик.
Активный транспорт - происходит по челночному механизму. Белок-переносчик на внешней стороне мембраны присоединяет молекулу субстрата и макроэргическую связь молекулы АТФ (комплекс: белок-переносчик – макроэргическая связь - субстрат). За счёт энергии связи комплекс перемещается на внутреннюю сторону мембраны, где распадается. Свободный переносчик возвращается на внешнюю сторону, которая имеет высокое химическое сродство (рис. 21).
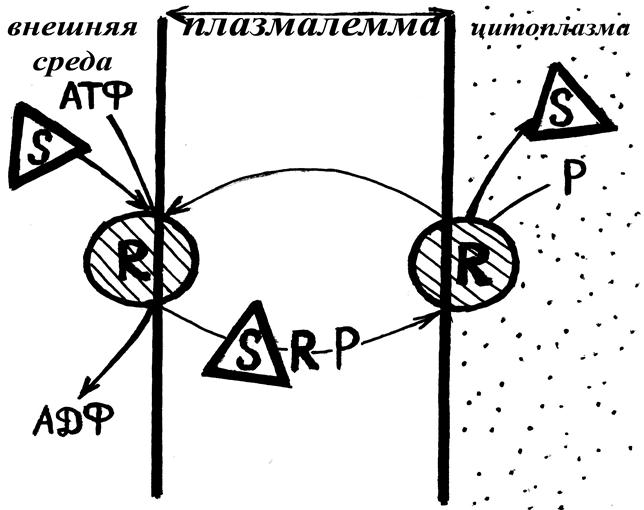
Рис.21. Схема активного поступления веществ в клетку через плазмалемму. S – субстрат; R – белок-переносчик; P – остаток фосфорной кислоты после гидролиза макроэргической связи.
15. Общая характеристика обмена веществ микроорганизмов
Обмен веществ – Метаболизм – совокупность всех химических реакций в клетке (организме).
Реакции метаболизма идут в двух направлениях:
Диссимиляция - (катаболизм)-совокупность всех реакций распада. На катаболическом направлении: 1). Из сложных веществ образуются простые - (белки расщепляются до аминокислот, жиры до глицерина и жирных кислот, крахмал до глюкозы, нуклеиновые кислоты до нуклеотидов. Далее мономеры расщепляются до СО2 и Н2О. 2). Реакции идут с выделением энергии |
Ассимиляция - (анаболизм) совокупность всех реакций синтеза. На Анаболическом направлении: 1). Из простых веществ образуются сложные - (из глюкозы синтезируются крахмал, гликоген, из глицерина и жирных кислот – жиры, из аминокислот – белки. При фотосинтезе из СО2 и Н2О образуются мономеры, например, глюкоза). 2)Реакции идут с поглощением энергии |
На катаболическом направлении образуются структурные блоки, из которых при ассимиляции образуются сложные вещества. При распаде веществ выделяется энергия, которая консервируется в виде макроэргической связи АТФ.
Типы углеродного питания микроорганизмов
Углерод – основной элемент питания всех живых организмов. Из углерода состоят углеродные цепи – основа всех органических веществ.
Способы углеродного питания микроорганизмов определяются в зависимости от источника углерода, источника водорода (или электронов) для восстановления, источника энергии для синтеза.
По источнику углерода все микроорганизмы подразделяются на автотрофные (получающие углерод путем усвоения СО2 из воздуха) и гетеротрофные (которые усваивают готовые органические вещества) (табл.3).
Таблица 3. Классификация способов углеродного питания микроорганизмов
|
Автотрофы Организмы, которые используют минеральные формы углерода (СО2) и образуют из них органические вещества |
Гетеротрофы Организмы, которые усваивают готовые органические вещества, перестраивая их на вещества своего тела |
||
|
И С Т О Ч Н И К У Г Л Е Р О Д А |
|||
|
СО2 |
Органические вещества |
||
|
ФОТОАВТОТРОФЫ |
ХЕМОАВТОТРОФЫ |
САПРОТРОФЫ |
ПАРАТРОФЫ |
|
Для синтеза органических веществ используют лучистую энергию солнца |
Для синтеза органических веществ используют энергию окисления минеральных веществ |
Используют готовые органические вещества мертвых тел |
Используют готовые органические вещества живых тел |
|
И с т о ч н и к э н е р г и и – |
|||
|
Лучистая энергия солнца. Работают по схеме: 6СО2+6Н2О®С6Н12О6+6О2 СО2+Н2S®С6Н12О6+H2SO4 |
Энергия окисления минеральных веществ: NH3 + О2 ® НNO3 + Е H2S + O2® H2SO4 + Е |
Энергия окисления органических веществ - дыхания и брожения: С6Н12О6+6О2®6СО2+6Н2О + Е С6Н12О6®2СН3СН2ОН + 2СО2 +Е |
|
|
П Р Е Д С Т А В И Т Е Л И |
|||
|
Водоросли -Ulothrix, Цианобактерии - Nostoc, Anabaena, Gloeocapsa, окрашенные серобактерии - Chlorobium, Chromatium |
Нитрифицирующие бактерии (Nitrobacter), Сульфофицирующие бактерии (Thiobacillus) |
Дрожжи: Saccharomyces vini, Уксусно-кислые бактерии - Acetobacter |
Salmonella, Shigella, Bdellovibrio |
По источнику энергии выделяют два основных типа метаболизма:
· Фототрофные Микроорганизмы, использующие в качестве источника энергии видимое электромагнитное излучение – свет;
· Хемотрофные микроорганизмы, получающие энергию в результате окисления различных субстратов (минеральных и органических).
По источнику водорода микроорганизмы подразделяются на:
· Метатрофные, использующие для восстановления электроны неорганических веществ (Н2, Н2S, NH3 , Fe++);
· Органотрофные, для которых донорами водорода и электронов служат органические соединения.
17. Фотосинтез у бактерий
Фотосинтез – процесс образования органических веществ из диоксида углерода (CO2) за счет лучистой энергии солнца (Hν). Это способ автотрофного углеродного питания. Различают два основных типа фотосинтеза у бактерий:
1. Оксигенный – Происходит с выделением кислорода по уравнению:
Hν
![]() 6CO2 + 6H2O C6H12 O6 + 6O2
6CO2 + 6H2O C6H12 O6 + 6O2
Возбудители этого процесса– цианобактерии (NostОс, Anabaena, Gloeocapsa).
2. Аноксигенный – Процесс не сопровождается выделением кислорода, идет по схеме:
Hν
![]() 6СO2 + 6H2S C6H12O6+ 6S
6СO2 + 6H2S C6H12O6+ 6S
Или:
![]() СO2 + S + H2O Hν C6H12O6 + Н2SO4
СO2 + S + H2O Hν C6H12O6 + Н2SO4
Возбудители – окрашенные серобактерии: зелёные – Chlorobium и пурпурные – Chromatium.
Хемосинтез у бактерий
Хемосинтезом называется синтез органических веществ из диоксида углерода (CO2) за счет энергии окисления минеральных веществ, таких как Н2, Н2S, S, NH3 , Fe++ и др. Это способ автотрофного усвоения углерода. Процесс идет по схеме:
+Е
![]() СO2 + H2S C6 H12 O6 + S
СO2 + H2S C6 H12 O6 + S
Источниками энергии для хемосинтеза являются следующие процессы:
1. Нитрификация: NH3 + O2→ HNO3 + E (возбудители – бактерии рода Nitrobacter);
2. Сульфофикация: H2S + O2 → H2SO4 + E (возбудители – бактерии рода Thiobacillus);
![]() 3. Окисление железа: Fe2+ —ē FЕ3+ + E (возбудители – бактерии рода GallionellА).
3. Окисление железа: Fe2+ —ē FЕ3+ + E (возбудители – бактерии рода GallionellА).
Азотное питание микроорганизмов
Азот необходим микробной клетке для синтеза белков, нуклеиновых кислот, полимеров клеточной стенки (муреина). В качестве источников азота микроорганизмы могут усваивать как минеральные, так и органические вещества.
По использованию источников азота
Микроорганизмы подразделяют на группы:
Аминоавтотрофы - Аминогетеротрофы -
Используют минеральные используют органические
формы азота: Формы азота:
- аммиак NH3 (или NH4+) белки, аминокислоты,
- нитратная форма NO3‾ нуклеотиды,
- молекулярный азот – N2 нуклеиновые кислоты
ПРИМЕРЫ:
Дрожжи, плесневые грибы, молочнокислые бактерии,
Уксусно-кислые бактерии гнилостные бактерии
Отношение микроорганизмов к источникам кислорода
Кислород является основным окислителем в метаболических процессах. Микроорганизмы могут его усваивать в свободном и связанном виде. Некоторые виды микробов могут существовать только в кислородной среде, для других кислород токсичен. Причиной этого является накопление в кислородной среде перекисей, которые очень токсичны для бактериальной клетки. У аэробов перекиси разрушает фермент каталаза, у строгих анаэробов этот фермент отсутствует.
Выделяют следующие группы микроорганизмов по отношению к источникам кислорода
Аэробы Анаэробы
Облигатные факультативные ОБлигатные
Облигатные (обязательные) аэробы – организмы, для жизнедеятельности которых необходим кислород воздуха. Подразделяются на 2 подгруппы:
1. Строгие облигатные аэробы нуждаются в концентрации кислорода, близкой к атмосферной (около 20%). Примером строгих облигатных аэробов могут служить уксусно-кислые бактерии (род Acetobacter);
2. МИкроаэрофилы Предпочитают низкие концентрации кислорода - до 10% (сальмонелла, бруцелла).
Облигатные анаэробы в кислороде не нуждаются. Среди них также выделяют 2 подгруппы:
1. СТрогие облигатные анаэробы В кислородной среде погибают. У них отсутствует фермент каталаза, разрушающий перекись водорода. Представителями этой группы являются бактерии рода Clostridium.
2. АЭротолерантные Микроорганизмы в кислороде не нуждаются, но в кислородной среде не погибают (например, молочно-кислые бактерии) .
Факультативные аэробы (они же факультативные анаэробы) могут переключаться с аэробного метаболизма на анаэробный. К ним относятся дрожжи рода Saccharomyces.
Брожение как способ получения энергии гетеротрофами
Гетеротрофы использую органические вещества и как строительный материал, и как источник энергии. Расщепляя ферментами углеводы в анаэробных условиях, микроорганизмы вызывают процесс брожения.
Брожение – Ступенчатый ферментативный анаэробный окислительно-восстановительный процесс, в котором микроорганизмы получают энергию, необходимую для жизнедеятельности. Рассмотрим схемы двух распространенных в природе процессов, названия которых даны по основным продуктам:
Молочнокислое брожение:
C6H12O6 → 2CH3CHOHCOOH + E
Молочная кислота
Спиртовое брожение:
C6H12O6 → 2CH3CH2OH + 2CO2 + E
С : О = 1 : 1 С : О = 2 : 1 С : О = 1 : 2
Этиловый спирт
На примере спиртового брожения убедительно доказывается, что брожение - окислительно-восстановительный процесс. Это хорошо видно по изменения соотношения углерода и кислорода в субстрате брожения (глюкоза C6H12O6 – соотношение С : О = 1 : 1) и продуктах (этиловый спирт С : О = 2 : 1; Углекислота - С : О = 1 : 2). Энергия в брожениях выделяется за счет перераспределения окислительных эквивалентов (кислорода) между продуктами спиртового брожения.
Характеристика брожения:
1.Субстраты брожения – органические вещества (углеводы, реже органические кислоты).
2.Продукты брожения – органические вещества (кислоты, спирты).
3.Биологический смысл процесса – получение энергии
4.Условия процесса – анаэробные (процесс идет без доступа кислорода)
5.Механизм брожения. Большинство брожений проходит в 2 этапа:
Первый этап - Универсальный (Чаще Гликолиз) – проходит по схеме:
C6 H12 O6 → 2CH3COСОOH + 2НАД • H2 + 2АТФ
Пировиноградная кислота (ПВК)
Энергия, полученная на этом этапе консервируется в виде макроэргических связей АТФ путем субстратного фосфорилирования.
Второй этап – специфический - зависит от набора ферментов, у разных брожений протекает по-разному. Если микроорганизм образует активный фермент ПВК-декарбоксилазу, то происходит декарбоксилирование (отщепление СО2) от пировиноградной кислоты с образованием уксусного альдегида, который затем восстанавливается водородом НАД•Н2 до этанола – схема спиртового брожения. Если ПВК-декарбоксилаза отсутствует, то пировиноградная кислота сразу восстанавливается до молочной водородом НАД•Н2, Которая образовалась на этапе гликолиза:
CH3CHOНCOOH
НАД•Н2 Молочная кислота
![]()
![]() CH3COCOOH фермент ПВК-декарбоксилаза СН3СНО НАД•Н2 СН3СН2ОН
CH3COCOOH фермент ПВК-декарбоксилаза СН3СНО НАД•Н2 СН3СН2ОН
ПВК Ацетальдегид Этанол
Таким образом, акцепторами водорода и электронов являются продукты самого брожения – органические вещества.
В результате процесса брожения высвобождается и используется для жизнедеятельности микроорганизма лишь часть энергии субстрата. Основная масса химической энергии остается в продуктах брожения – спиртах, кислотах. Объясняется неполное высвобождение энергии субстрата при брожении отсутствием внешнего окислителя, в первую очередь, кислорода.