Печень
Печень млекопитающих определяется, как полифункциональный орган. На фоне главной функции в составе аппарата пищеварения, она выполняет десятки других, предопределенных, прежде всего, активным ее участием в метаболических реакциях организма. У новорожденных животных ткани печени содержат значительное количество биологически активных веществ, которые влияют на весь организм (табл. 97).
Таблица 97
Количество биологически активных веществ в печенке
Неонатальных телят (Рой Дж. Х. Б., 1973)
|
№ п/п |
Показатель |
Значение |
Авторы |
|
1. |
Содержание меди А) на 1кг живой массы, мг Б) на 1кг сухого вещества печени, мг |
14 100 |
Rusoff L. L., 1941 |
|
2. |
Содержание витамину А в 100 г печени, мкг |
75 |
Walker D. M. et al., 1949 |
|
3. |
Содержание в 1г печени (в дужках – колебание), в мкг: А) биоплетень Б) фоллиевая кислота В) никотиновая кислота Г) пантотеновая кислота Д) рибофлавин Е) тиомин Ж) витамин В6 (пирдоксин) З) витамин В12 |
0,095 (0,08-0,12) 0,7 (0,3-0,9) 74 (54-86) 18 (16-22) 9 (8-10) 1,2 (1,0-1,3) 1,9 (1,8-2,0) 0,25 (0,15-0,36) |
Ford J. E., |
В учебной и научной литературе печень считают “биохимической лабораторией” или “метаболическим мозгом” организма. Полифункциональнисть печени отражает определенные особенности ее строения и топографии, которые являются характерными для представителей разных видов млекопитающих, в том числе и домашных [6, 243, 348]. У половозрелых домашных животных, печень является наиболее большой застенной железой органов пищеварения. У крупного рогатого скота она достигает относительной массы 1,1%, у свиней 2,5% и собак 2,8-3,4% живой массы. При этом у всех животных печень разделяется на доли– левую, среднюю (хвостатую и квадратную) и правую. У всеядных и хищных животных ее левая и правая доли дополнительно разделяются еще на латеральную и медиальную. Расположен орган в краниальном участке эпигастрия и прикреплен связками к диафрагме.
Печень новорожденных млекопитающих проявляет лишь некоторые дефинитивные признаки строения, что отражает ее подобие с взрослыми животными (деление ее на доли). Относительная масса печени суточных телят составляет 2,41-3,21%, у поросят – 2,07-3,96% и щенков – 4,55-5,98% живой массы, которая предопределяет ее значительные размеры и соответствующий объем, который она занимает в брюшной полости [164]. В течение следующих 10 и 20 суток постнатальной жизни животных относительная масса печени уменьшается: у телят до 2,11-2,45% и 2,23-2,62%, у поросят – 3,36-4,43% и 3,76-4,94% и щенков – 3,80-5,22% и 4,36-4,99%, что свидетельствует о начале становления дефинитивных функций и в процессах пищеварения [73, 291]. Такое уменьшение относительной массы печени является проявлением общебиологической закономерности динамики массы органа, как в пре-, так и в постнатальном онтогенезе (табл. 98).
Таблица 98
Динамика абсолютной и относительной массы печени у домашных животных в онтогенезе (А. С. Чалый, 1966)
|
Возраст месяцы |
Крупный рогатый скот |
Овцы |
Свиньи |
|||
|
Абсолютная |
Относительная |
Абсолютная |
Относительная |
Абсолютная |
Относительная |
|
|
Пренатальный |
– |
– |
– |
– |
0,82 |
13,9 |
|
1,5-2 |
0,51 |
5,5 |
0,51 |
8,5 |
2,36 |
7,6 |
|
2-3 |
8,70 |
4,3 |
4,2 |
6,5 |
6,6 |
4,67 |
|
3-4 |
22,3 |
3,9 |
29,1 |
6,2 |
17,1 |
2,13 |
|
4-5 |
72,0 |
3,7 |
55,3 |
3,3 |
– |
– |
|
5-6 |
172,4 |
3,1 |
– |
– |
– |
– |
|
6-7 |
252,5 |
2,4 |
– |
– |
– |
– |
|
7-8 |
460,0 |
2,3 |
– |
– |
– |
– |
|
Новорожденные |
641,7 |
2,1 |
– |
– |
– |
– |
|
до 2 мес |
– |
– |
71,8 |
1,7 |
– |
– |
|
2-4мес |
– |
– |
– |
– |
435,8 |
1,75 |
|
1-3 года |
2848,0 |
1,35 |
– |
– |
– |
– |
|
до 2 лет |
– |
– |
483,7 |
1,4 |
1126,0 |
1,44 |
|
до 5 лет |
– |
– |
564,9 |
1,1 |
– |
– |
|
4-14 лет |
5054,0 |
1,27 |
– |
– |
– |
– |
Динамика микроскопических изменений в печенке протекает на фоне существенных трансформаций интраорганных тканевых структур и кровеносных сосудов, что характерно для онтогенеза млекопитающих [318].
Гистогенез печени у млекопитающих начинается на 22-28 день внутриутробного развития [13, 27, 34, 55, 374]. Она формируется в виде печеночного дивертикула вентральной стенки первичной кишки. У зародышей крупного рогатого скота паренхима органа образована гепатоцитами, между которыми размещены широкие синусоиды. При этом, гепатоцити имеют форму продолговатых пластинок толщиной в 2-5 клеток. Афферентными и эфферентными кровеносными сосудами печени в этот период являются ветви желточно-брыжеечных вен.
В предплодный период развития проявляется трубчатый характер структуры паренхимы печени, при котором желчные капилляры образованы 2-4 клетками. До конца предплодного и начала плодного периодов (3-5 месяцев) в печени активно проявляется гемопоэз (гепатолиенальный период). На начальных стадиях он проходит эндоваскулярно, подобно с ранним гемопоэзом в мезенхиме желточного пузыря, и изменяется экстраваскулярной стадией.
Главными афферентными сосудами являются пупочные вены, и больше – левая. Печеночная артерия, в частности у предплодов крупного рогатого скота, при более поздней закладке (на 38 сутки), в течение постнатального онтогенеза является сосудом с малым поперечником. Эфферентными сосудами в предплодный период развития являются печеночные вены, которые сообщается с каудальной полой веной.
В плодный период, к родам, тяжи гепатоцитов становятся тоньше, в паренхиме растет численность балок и уменьшается поперечник синусоидов. Гемопоэз в печени в это время затухает. Первые признаки печеночных долек, у плодов крупного рогатого скота, возникают после 4 месяца пренатального онтогенеза. Однако доли дифференцируются гистологически лишь после 7-8 месячного развития плодов. Завершается их формирование у 6-9-месячных животных после рождения.
Особенного внимания заслуживает наличие центров гемопоэза в печени новорожденных млекопитающих. Если у суточных телят центры гемопоэза не определяются, то у животных с менее выраженной матуронатностью организма они наблюдаются еще некоторое время после рождения. У новорожденных поросят центры гемопоэза обнаруживаются в первые дни постнатального периода онтогенеза. У имматуронатных животных (крыс) они еще сохраняются в течение нескольких недель.
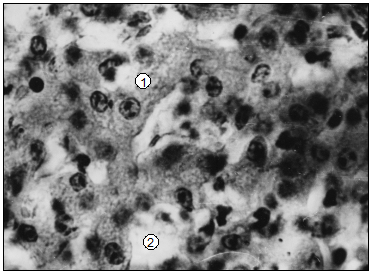
Наши исследования показывают, что у новорожденных домашных животных паренхима печени имеет определенные черты незавершенности строения [174, 200]. У суточных телят гепатоцити имеют круглую форму. Их цитоплазма – мелкозерниста (рис. 90, А). Структура в разных частицах печени гепатоцитов подобна. В дольках количество гепатоцитов непостоянно и изменяется в течение первых 20 суток постнатальной жизни животного (табл. 99). У телят суточного возраста количество гепатоцитов меньше сравнительно из 10- и 20-суточными животными. При этом их количество преобладает в левой доле. Доли паренхимы печени суточных телят имеют незавершенную структуру. Граница выраженна не четко и может отсутствовать с одной или разных сторон доли, которая предоставляет группам долек, размещенных близко друг возле друга, в виде конгломератов. Центральные участки таких печеночных долек сформированы соединением радиальных балок, а перипортальные содержат паренхиму губчатой структуры. Паренхима размещена в субкапсулярной зоне долек печени, имеет подобную стуктуру. Ячейки гемопоэза в паренхиме печени у суточных телят отсутствуют.
У 10-суточных телят, количество гепатоцитов увеличивается, тогда как в 20 – несколько уменьшается (при росте их в левой доле). Следует отметить, что печень телят 10- и 20-суточного возраста содержит незавершенной структуры доли, которые более четко отграничиваются одна от другой, сравнительно с суточными животными. Губчатой структуры паренхимы в частицах печени почти не оказывается.
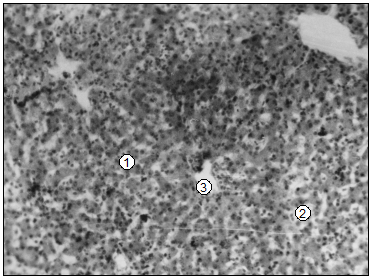
У новорожденных (суточных) поросят паренхима печени имеет еще более незавершенную структуру сравнительно с телятами: гепатоцити круглые и содержат цитоплазму с мелкой зернистостью, которая заполняет всю клетку (рис.90, Бы). Встречаются отдельные гепатоцити, которые имеют прозрачную и, даже, оптически опустошенную цитоплазму. Такого типа клетки преобладают у суточных поросят с низкой (500-600 г) живой массой. Количество гепатоцитов у суточных поросят больше, чем у телят. В долях печени оно неодинаково, и несколько преобладает в средней доле (табл. 100). Дольки в печени суточных поросят не обнаруживаются. Гепатоцити образуют разветвленные балки и скопления из 2-4 клеток, которые размещаются между афферентными и эфферентными сосудами в виде густой “сетки”, которая придает паренхиме губчатый вид. Кроме того, в печени суточных поросят среди гепатоцитов определяются центры гемопоэза в виде отделенных островков (табл. 101). Их количество очень непостоянно и преобладает у поросят с низкой живой массой.
У 10-суточных поросят в центральных участках долек печени, находятся не четко ограниченные доли. При этом количество гепатоцитов значительно уменьшается, особенно в средней доле печени, а мелкие ячейки гемопоэза становятся одиночными. С возрастом поросят (20-суточных), печень достигает определенной структуры, в которой оказываются доли, которые четко отграничены одна от другой тонкими прослойками рыхлой волокнистой соединительной ткани, а балки гепатоцитов приобретают радиальное размещение. Количество гепатоцитов в дольках растет, и больше в средней доле печени.
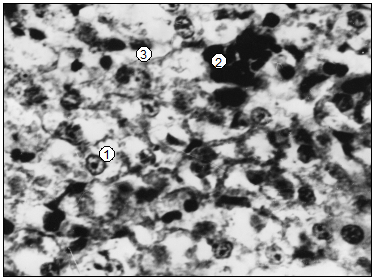
У суточных щенков во всех долях печени находится значительное количество оптически опустошенных гепатоцитов (рис. 91). Клетки, с окрашенной цитоплазмой, образуют островки без четких границ в средних участках долек, а также в субкапсулярной зоне печени. Количество гепатоцитов у щенков меньше, сравнительно с печенью поросят и телят. Печеночных долек у суточных щенков не проявляется, гепатоцити формируют скопления гомогенной структуры, в которых без четкой ориентации проходят синусоидные капилляры. Центры гемопоэза у суточных щенков имеют относительную площадь уже почти в 2-4 раза большую, чем у поросят, на фоне значительно меньшей вариабельности (табл. 102).
I II
А )

I II
В)
Рис. 90. Гистотопограма печени телят – А, поросят – В (сутки). Гематоксилин и эозин, МБИ-6. I – 10х9. II – 10х40 (В. В. Лемещенко): 1 – гепатоцити; 2 – синусоиды; 3 – центры гемопоэза.
I II
Рис. 91. Гистотопограммы печени щенят (сутки). Гематоксилин и эозин МБИ-6. I – 10х9. II – 10х40. 1. Гепатоцити. 2. Центры гемопоэза (В. В. Лемещенко).
Таблица 99.
Динамика количества гепатоцитов в 1мм 2
в долях печени 1-, 10- и 20-суточных телят (В. В. Лемещенко)
|
Возраст, |
Доли печени |
Всего по |
||
|
Левая |
Середня* |
Правая |
||
|
1 |
4822,22±351,19 |
4444,44±94,96 |
4369,66±244,33 |
4433,76±112,4 |
|
V,% |
12,61 |
3,71 |
9,68 |
4,39 |
|
10 |
3523,51±678,99 |
5170,92±352,22 |
5181,62±259,99 |
5290,60±415,45 |
|
V,% |
21,29 |
11,80 |
8,67 |
13,60 |
|
20 |
5220,08±95,17 |
4882,48±661,10 |
5192,30±218,17 |
5098,29±315,39 |
|
V,% |
3,16 |
23,45 |
7,28 |
10,71 |
Таблица 100
Динамика количества гепатоцитов в 1 мм2 гистотопограммы долек печени 1-, 10- и 20-суточных поросят (В. В. Лемещенко)
|
Возраст, |
Доли печени |
Всего по |
||
|
Левая* |
Средняя |
Правая* |
||
|
1 |
5370,19±272,87 |
5439,10±557,26 |
5260,68±340,04 |
5335,83±328,87 |
|
V,% |
12,45 |
25,10 |
15,84 |
15,10 |
|
10 |
4246,80±178,69 |
3798,08±115,31 |
4384,62±277,09 |
4143,16±176,33 |
|
V,% |
8,42 |
6,08 |
12,64 |
8,51 |
|
20 |
4895,30±652,91 |
5741,43±756,16 |
4861,11±474,96 |
4832,62±446,14 |
|
V,% |
23,10 |
22,81 |
16,92 |
15,99 |
* – левая (правая) латеральная и медиальная доли
Таблица 101
Относительная площадь центров гемопоэза в долях печени суточных поросят и щенков (%) (В. В. Лемещенко)
|
Вид |
Доли печени |
Всего по |
||
|
Левая* |
Средняя |
Правая* |
||
|
Поросята |
1,13±0,92 |
0,71±0,69 |
0,56±0,32 |
0,80±0,63 |
|
V,% |
160,53 |
192,96 |
116,07 |
157,50 |
|
Щенки |
2,99±0,54 |
2,15±0,39 |
4,00±0,31 |
3,04±0,13 |
|
V,% |
35,79 |
36,28 |
15,50 |
8,15 |
· – то же, что в табл. 103
· Таблица 102
Динамика количества гепатоцитов в 1 ммб гистотопограмм
Печени 1-, 10- и 20-суточных щенков (В. В. Лемещенко)
|
Возраст, |
Доли печени |
Всего по |
||
|
Левая** |
Середняя* |
Правая** |
||
|
1 |
4070,51±632,36 |
3733,98±470,55 |
3900,64±372,96 |
3901,71±431,95 |
|
V,% |
31,07 |
25,20 |
19,21 |
22,14 |
|
10 |
4912,39±246,12 |
4816,24±209,40 |
4478,63±235,66 |
4735,75±146,00 |
|
V,% |
8,98 |
7,53 |
9,11 |
5,34 |
|
20 |
4262,82±126,10 |
4070,51±273,85 |
3756,41±314,65 |
4029,91±95,59 |
|
V,% |
5,12 |
11,65 |
14,51 |
4,11 |
* но ** – те же, что в табл. 95
У 10-суточных щенков все гепатоцити имеют окрашенную цитоплазму и растут в количестве, особенно в левой и средней долях. Их количество уменьшается больше в правой доле в 20-суточном возрасте. При этом доли в печени у щенков не определяются.
Следует отметить о значительной вариабельности количества гепатоцитов у суточных домашных животных, которая уменьшается с формированием долей в паренхиме печени. До конца новорожденного периода ваиабельность количества гепатоцитов составляет 4-15%.
Особенности внутреннего строения печени домашных новорожденных животных предопределены наличием в ней провизорных структур, что вызвано активной динамикой функций органа. Изменение функциональной активности печени вызывается, в первую очередь, адаптогенезом ее кровеносных сосудов, которые в этот период жизни животных определяют объем крови, которая будет оттекать оторганов брюшной полости к сердцу.
Афферентная кровеносная система печени млекопитающих состоит из интраорганых разветвлений воротной вены и печеночной артерии [24, 197, 348, 378]. При этом, около 75-80% кислорода и питательных веществ поступает в печень из крови воротной вены. Кровь выносится многочисленными печеночными венами, которые впадают в каудальную полую вену [198, 200].
У взрослых млекопитающих соединения портальной и кавальной кровеносных систем осуществляется через синусоидную сетку печени. У новорожденных еще несколько суток после рождения (у телят в условиях гиподинамии – до 20-10 дней), функционирует венозный проток, который считают продолжением пупочной (афферентной вены) к каудальной полой вене. У новорожденных телят венозный проток обеспечивает течение крови к 5-20-суточному возрасту, у щенков – 5-10. Характерно, что у поросят, она облитерируется значительно раньше. Облитерация проходит еще до рождению или через двое суток после рождения и остается в виде разветвленных анастомозов [242,375, 376].
Как показывают наши исследования, кровоток в печени новорожденных телят, поросят и щенков связана с активностью афферентного (пупочных ворот) и эфферентного (кавального) коллекторов (рис. 92, 93). Афферентный коллектор печени животных образуется к их рождению путем анастомоза соответствующих вен в воротах печени. У суточных млекопитающих пупочная вена в брюшной полости имеет просвет, заполненный кровью. В участке ворот печени она соединяется с воротной веной и, от сформированного, таким образом, коллектора, ответвляются интраорганнные ветви, а также венозный проток. Интраорганные ветви коллектора пупочных ворот в сопровождении более тонких ветвей печеночной артерии, образуют паравенозное сплетение и формируют лобарные афферентные магистрали.
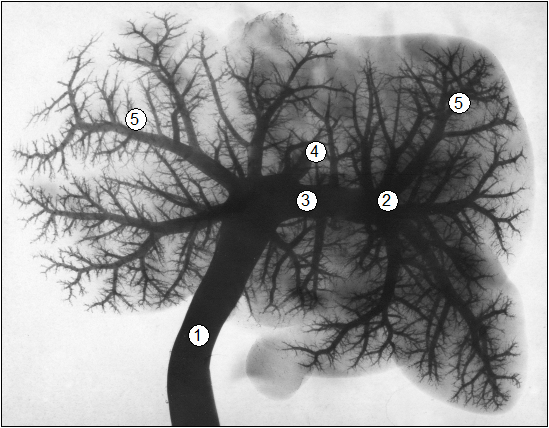
Рис. 92. Рентгенограмма афферентных вен печени суточного теленка (сутки) (В. В. Лемещенко): 1 – пупочная вена; 2 – воротная вена; 3 – афферентний коллектор; 4 – венозный проток; 5 – внутриорганные ветви.
Эфферентний коллектор печени у новорожденных животных сформирован каудальной полой веной, которая проходит в желобе или канале вдоль тупого края органа и принимает, соответственно, внутриорганные магистрали (печеночные вены) и венозный проток.
Венозный проток у домашных суточных животных имеет видовые особенности (рис. 94). У телят он является магистральным, слабо выгнутым сосудом, который проходит под тонким слоем паренхимы и не имеет боковых ветвей. Начинается суженным концом с левой стороны афферентного коллектора и впадает расширенным – в каудальную полую вену рядом с левой печеночной.
У суточных поросят венозный проток магистрального типа отсутствует. С левой стороны пупочных ворот коллектора разветвляется 3-6 древовидных вен, ветви которых проходят как в паренхиме, на границе, левой латеральной и хвостатой долей, так и впадают к боковому, расширенному разветвлению левой печеночной вены. Следовательно, у поросят образуются макроскопические множественные порто-кавальные анастомозы.
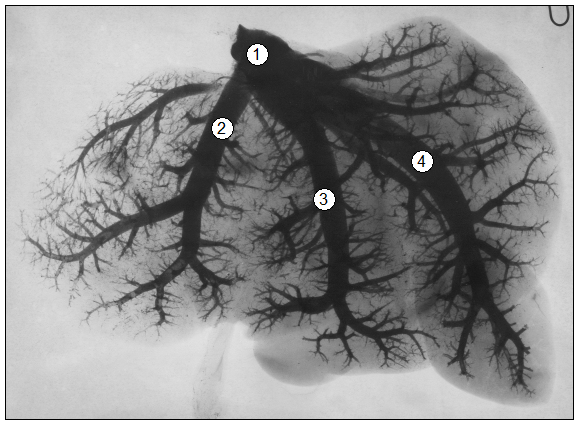
Рис. 93. Рентгенограмма вен печени теленка (сутки): 1 – кавальний коллектор 2 – левая печеночная вена; 3 – средняя печеночная вена; 4 – вена хвостатого отростка (В. В. Лемещенко).
У суточных щенков венозный проток тонкий, прямой, не разветвляется и почти не прикрыт паренхимой печени (лишь в незначительном участке, каудальнее диафрагмы). Перед впадением в каудальную полую вену он ампулообразно расширяется и принимает левую диафрагмальную вену (Lacus venae cavae).
У поросят и, особенно, щенков 10-суточного возраста пупочная вена облитерируется (только в одиночных случаях сохраняется прерывистый, извилистый канал), как и венозный проток (множественные анастомозы), а коллектор пупочных ворот начинает превращаться в левую ветвь воротной вены.
У 10-суточных телят, на фоне уменьшения размеров пупочной вены и венозного протока, просветы содержат кровь. В 20-суточном возрасте телят венозный проток при нормальном статусе организма, как правило, облитерируется, при сохранении незначительного просвета в пупочной вене, коллектор пупочных ворот, также, как у поросят и щенков, превращается в левую ветвь воротной вены.
Динамика площади просвета и стенки афферентных кровеносных сосудов печени домашных животных в первые 20 суток постнатальной жизни происходит асинхронно.
В суточном возрасте у животных наибольший просвет, среди афферентных сосудов имеет воротная вена (табл. 103-105). Однако площадь стенки существенно преобладает в пупочной вене, а коллектор пупочных ворот имеет промежуточные значения. При этом площадь просвета и стенки печеночной артерии у телят, поросят и щенков значительно больше малых, чем афферентных вен, подтверждая, что в суточном возрасте у животных основной объем крови печени притекает по воротной вене при наличии еще провизорной пупочной вены.
В 10-суточном возрасте новорожденных динамика показателей площади просвета и стенки пупочной вены не равнозначная. У телят она уменьшается, а у поросят – растет. У щенков, почти на всем протяжении пупочной вены, от пупочного кольца до афферентного коллектора, просвет пупочной вены не дифференцируется. Вместо него оказываются извилистые, прирывчастые, разветвленные каналы, которые свидетельствуют о ее облитерации. У 10-суточных телят в воротной вене увеличивается, как просвет, так и толщина стенки, у поросят они несколько уменьшаются, а у щенков – при утолщении стенки, уменьшается просвет. Параметры афферентного коллектора у поросят увеличиваются, у щенков-уменьшаются, а у телят – изменяются асинхронно. Такие же незначительные асинхронные изменения проявляются в печеночной артерии. До 10-х суток после рождения, у животных проявляется тенденция к трансформации пупочных ворот коллектора в левую ветвь воротной вены при активной перестройке (к полной облитерации у щенков) пупочной вены.
У 20-суточных поросят, как и у щенков, происходит облитерация пупочной вены. У телят, при несколько увеличенной площади ее стенки, значительно уменьшается просвет. Параметры воротной вены у животных растут, а в коллекторе пупочных ворот изменяются асинхронно. В печеночной артерии, при росте площади стенки вен, просветы почти не изменяется или несколько уменьшается. Такая динамика площади просвета и стенки афферентных сосудов печени у животных указывает на то, что течение и перераспределение крови происходит преимущественно воротной веной. Печеночная артерия с возрастом животных еще меньше играет роль в афферентном кровообращении печени. Характерно, что на фоне асинхронной динамики площади просвета и толщины стенки афферентных сосудов печени у животных 20-суточного возраста, вариабельность этих параметров колеблется в пределах средных значений.
Эфферентний коллектор суточных млекопитающих образуется каудальной полой веной. Перед печенью каудальная полая вена принимает кровь от тазовых конечностей, стенок и парных органов брюшной полости. От воротной вены, непосредственно через венозный проток и “чудесную сеть ” органа, и дальше – в печеночные вены. Эфферентный коллектор принимает кровь от объемных непарных органов брюшной полости. Вследствие этого он является структурой, которая регулирует интенсивность кровотока через диафрагму в грудную полость, то есть к сердцу.
Таблица 103
Динамика площади просвета и стенки афферентных кровеносных сосудов печени 1-, 10- и 20-суточных телят, мм2 (В. В. Лемещенко)
|
Возраст, Суток |
Показатели |
Кровеносные сосуды |
|||
|
Пупочная вена |
Воротная вена |
Коллектор пупочных ворот |
Печеночная артерия |
||
|
1 |
Просвет |
6,15±1,30 |
23,55±5,21 |
17,58±9,32 |
1,57±0,22 |
|
V,% |
47,48 |
49,47 |
91,57 |
31,65 |
|
|
Стенка |
18,98±3,18 |
29,88±4,31 |
12,60±3,45 |
1,92±0,43 |
|
|
V,% |
37,40 |
32,30 |
47,46 |
49,22 |
|
|
10 |
Просвет |
0,66±0,13 |
35,37±0,88 |
24,61±15,48 |
1,54±0,25 |
|
V,% |
33,33 |
4,32 |
108,98 |
28,57 |
|
|
Стенка |
9,89±2,03 |
31,64±2,21 |
9,96±2,55 |
3,95±0,89 |
|
|
V,% |
35,59 |
12,41 |
44,38 |
38,76 |
|
|
20 |
Просвет |
1,32±0,67 |
43,95±3,33 |
41,75±10,40 |
1,54±0,34 |
|
V,% |
87,78 |
13,13 |
43,16 |
37,66 |
|
|
Стенка |
12,81±3,57 |
29,95±3,67 |
16,70±1,52 |
2,71±0,84 |
|
|
V,% |
48,80 |
23,60 |
15,80 |
53,51 |
Таблица 104
Динамика площади просвета и стенки афферентных кровеносных сосудов печени 1-, 10- и 20-суточных поросят, ммб (В. В. Лемещенко)
|
Возраст Суток |
Показатели |
Кровеносные сосуды |
|||
|
Пупочная вена |
Воротная вена |
Коллектор пупочных ворот |
Печеночная артерия |
||
|
1 |
Просвет |
0,05±0,002 |
1,43±0,15 |
0,99±0,19 |
0,10±0,01 |
|
V,% |
25,00 |
23,08 |
38,10 |
10,00 |
|
|
Стенка |
0,74±0,11 |
1,06±0,19 |
0,72±0,10 |
0,46±0,15 |
|
|
V,% |
35,14 |
39,62 |
38,39 |
58,70 |
|
|
10 |
Просвет |
0,17±0,03 |
0,83±0,24 |
1,15±0,36 |
0,23±0,04 |
|
V,% |
47,06 |
63,86 |
70,43 |
34,78 |
|
|
Стенка |
0,99±0,19 |
0,98±0,22 |
0,92±0,20 |
0,46±0,08 |
|
|
V,% |
46,46 |
51,02 |
47,83 |
32,61 |
|
|
20 |
Просвет |
– |
2,09±0,38 |
1,71±0,28 |
0,14±0,04 |
|
V,% |
– |
35,88 |
32,16 |
57,14 |
|
|
Стенка |
– |
1,76±0,24 |
0,99±0,14 |
0,51±0,05 |
|
|
V,% |
– |
27,27 |
28,28 |
19,61 |
Характерно, что печеночные вены у суточных телят, поросят и щенков способных пропускать крови, больший объема, который протекает по каудальной полой вене. У суточных животных печеночные вены образуют более, чем 15-20 стволов разного поперечника, среди которых определяются три наиболее большие, которые впадают в эфферентный коллектор на всем его протяжении, равномерно распределяя интраорганные дренажные токи крови.
У суточных животных (как и у 10- и 20-суточных) суммарная площадь просвета и стенки печеночных вен значительно больше, чем афферентного коллектора (табл. 106-108). Характерно, что лишь у телят площадь торакальной части коллектора меньше абдоминальной. У поросят и щенков проявляется обратная тенденция. В 10-суточном возрасте животных абдоминальная часть кавального коллектора печени растет: у телят больше за счет площади стенки, а у поросят – просвета. Тогда как у щенков площадь просвета даже уменьшается. Подобная закономерность наблюдается и в торакальной части эфферентного коллектора, параметры которого уменьшаются у щенков. При этом, площадь просвета, толщины и стенки печеночных вен уменьшается как у щенков, так и у телят.
У животных 20-суточного возраста проявляется тенденция к уменьшению площади печеночных вен. Параметры Эфферентного коллектора (за исключением поросят) растут, а их вариабельность не превышает средных значений [196, 199, 200].
Следовательно, динамика площади просвета и стенки кровеносных сосудов печени свидетельствует, что у животных к 20-суточному возрасту афферентный коллектор трансформируется в левую ветвь воротной вены при уменьшении параметров дренажных печеночных вен. Асинхронная динамика площади просвета и стенки эфферентного коллектора указывает на непостоянный характер интенсивности тока крови по нему с органов брюшной полости до сердца.
Таблица 105
Динамика площади просвета и стенки афферентных кровеносных сосудов печени 1-, 10- и 20-суточных щенков, мм2 (В. В. Лемещенко)
|
Возрас, суток |
Показатели |
Кровеносные сосуды |
|||
|
Пупочная вена |
Воротная вена |
Коллектор пупочных ворот |
Печеночная артерия |
||
|
1 |
Просвет |
0,24±0,07 |
0,88±0,44 |
0,65±0,15 |
0,18±0,07 |
|
V,% |
58,33 |
87,50 |
38,46 |
77,78 |
|
|
Стенка |
0,70±0,12 |
0,43±0,15 |
0,36±0,11 |
0,36±0,10 |
|
|
V,% |
35,71 |
58,14 |
52,72 |
55,56 |
|
|
10 |
Просвет |
– |
0,53±0,18 |
0,42±0,01 |
0,13±0,01 |
|
V,% |
– |
66,04 |
7,14 |
15,38 |
|
|
Стенка |
– |
0,58±0,16 |
0,30±0,05 |
0,34±0,04 |
|
|
V,% |
– |
56,90 |
36,67 |
23,53 |
|
|
20 |
Просвет |
– |
1,07±0,21 |
0,38±0,16 |
0,13±0,01 |
|
V,% |
– |
39,25 |
73,68 |
15,38 |
|
|
Стенка |
– |
1,51±0,27 |
0,22±0,06 |
0,27±0,02 |
|
|
V,% |
– |
27,27 |
28,28 |
19,61 |
Таблица 106
Динамика площади просвета и стенки эфферентных вен печени 1-, 10- и 20-суточных телят, мм2 (В. В. Лемещенко)
|
Возраст, суток |
Показатели |
Кровеносные сосуды |
||
|
Абдоминальная часть |
Печеночные вены |
Торакальна часть |
||
|
1 |
Просвет |
38,14±12,31 |
145,16±14,92 |
33,75±4,06 |
|
V,% |
71,45 |
17,79 |
26,90 |
|
|
Стенка |
32,69±3,30 |
42,71±10,30 |
19,73±3,22 |
|
|
V,% |
22,56 |
41,77 |
36,49 |
|
|
10 |
Просвет |
46,58±6,09 |
83,50±7,20 |
58,30±5,33 |
|
V,% |
22,64 |
14,92 |
15,83 |
|
|
Стенка |
51,01±2,00 |
37,79±3,09 |
15,82±1,83 |
|
|
V,% |
6,80 |
14,16 |
20,03 |
|
|
20 |
Просвет |
75,59±8,12 |
79,55±0,25 |
67,30±16,45 |
|
V,% |
18,60 |
5,53 |
42,34 |
|
|
Стенка |
53,22±4,85 |
37,42±2,28 |
21,39±5,57 |
|
|
V,% |
15,78 |
10,58 |
46,75 |
Таблица 107
Динамика площади просвета и стенки эфферентных вен печени 1-, 10- и 20-суточных поросят, мм2 (В. В. Лемещенко)
|
Возрас, сутокт |
Показатели |
Кровеносные сосуды |
||
|
Абдоминальная часть |
Печеночные вены |
Торакальная часть |
||
|
1 |
Просвет |
1,87±0,60 |
11,35±1,38 |
6,15±0,79 |
|
V,% |
84,49 |
27,22 |
31,54 |
|
|
Стенка |
1,19±0,27 |
2,18±0,40 |
1,25±0,17 |
|
|
V,% |
60,50 |
38,70 |
32,80 |
|
|
10 |
Просвет |
5,01±1,33 |
14,48±1,06 |
6,37±1,73 |
|
V,% |
55,38 |
16,44 |
66,72 |
|
|
Стенка |
2,02±0,14 |
3,60±0,13 |
1,50±0,20 |
|
|
V,% |
17,33 |
8,06 |
32,67 |
|
|
20 |
Просвет |
4,34±0,23 |
14,35±0,64 |
7,94±2,54 |
|
V,% |
9,68 |
7,74 |
63,98 |
|
|
Стенка |
1,68±0,16 |
3,92±0,28 |
1,32±0,20 |
|
|
V,% |
19,05 |
15,53 |
29,55 |
Таблица 108
Динамика площади просвета и стенки эфферентных вен печени 1-, 10- и 20-суточных щенков, мм2 (В. В. Лемещенко)
|
Возраст Суток |
Показатели |
Кровеносные сосуды |
||
|
Абдоминальная часть |
Печеночные вены |
Торакальная часть |
||
|
1 |
Просвет |
1,78±0,48 |
14,94±1,02 |
1,15±0,21 |
|
V,% |
53,37 |
11,78 |
31,30 |
|
|
Стенка |
0,74±0,12 |
8,20±1,06 |
1,04±0,03 |
|
|
V,% |
33,78 |
22,32 |
4,81 |
|
|
10 |
Просвет |
0,57±0,14 |
10,82±0,89 |
0,69±0,13 |
|
V,% |
56,14 |
16,54 |
42,03 |
|
|
Стенка |
0,72±0,16 |
3,65±1,29 |
0,83±0,14 |
|
|
V,% |
50,00 |
70,68 |
37,35 |
|
|
20 |
Просвет |
1,90±0,44 |
8,92±2,84 |
1,48±0,30 |
|
V,% |
40,53 |
55,26 |
40,94 |
|
|
Стенка |
1,83±0,41 |
3,52±1,01 |
2,03±0,23 |
|
|
V,% |
38,80 |
50,00 |
22,06 |
|
Возраст Суток |
Показатели |
Кровеносные сосуды |
||
|
Абдоминальная |
Печеночные вены |
Торакальная часть эфферентного коллектора |
||
|
1 |
Просвет |
1,78±0,48 |
14,94±1,02 |
1,15±0,21 |
|
V,% |
53,37 |
11,78 |
31,30 |
|
|
Стенка |
0,74±0,12 |
8,20±1,06 |
1,04±0,03 |
|
|
V,% |
33,78 |
22,32 |
4,81 |
|
|
10 |
Просвет |
0,57±0,14 |
10,82±0,89 |
0,69±0,13 |
|
V,% |
56,14 |
16,54 |
42,03 |
|
|
Стенка |
0,72±0,16 |
3,65±1,29 |
0,83±0,14 |
|
|
V,% |
50,00 |
70,68 |
37,35 |
|
|
20 |
Просвет |
1,90±0,44 |
8,92±2,84 |
1,48±0,30 |
|
V,% |
40,53 |
55,26 |
40,94 |
|
|
Стенка |
1,83±0,41 |
3,52±1,01 |
2,03±0,23 |
|
|
V,% |
38,80 |
50,00 |
22,06 |
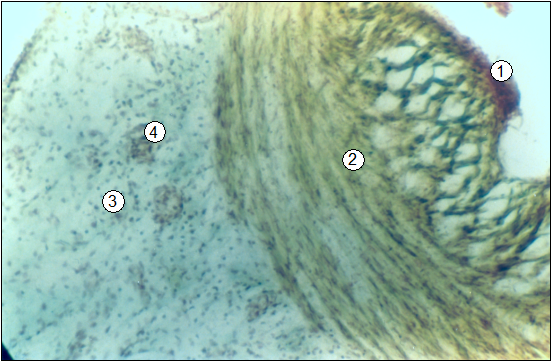
Рис. 95. Гистотопограма пупочной вены поросенка (сутки). Гематоксилин-пикроиндигокармин. МБИ-6, 10х8 (В. В. Лемещенко): 1 – внутренняя оболочка; 2 – средняя оболочка; 3 – внешняя оболочка и параумбиликальная соединительная ткань; 4 – параумбиликальные кровеносные сосуды.
Особенности архитектоники эктра - и интраорганных кровеносных магистралей печени новорожденных животных показывают, что рядом с незавершенностью их структуры, при наличии провизорных сосудистых компонентов (пупочной вены, венозного протока), уже у животных суточного возраста формируются дефинитивные признаки их строения. Наличие сформированных лобарных магистралей и коллекторов, которые перераспределяют потоки афферентной и эфферентной крови печени, указывают на интенсивное течение крови через печень у телят, поросят и щенков в первые 20 суток постнатальной жизни. Этот факт подтверждается и строением стенки кровеносных сосудов печени, особенно магистральных, в которой у суточных животных, определяются три оболочки с выраженными мускульными и волокнистыми компонентами.
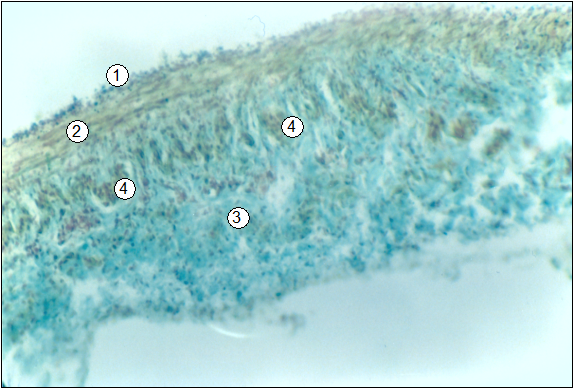
Рис. 96. Гистотопограма воротной вены поросенка (сутки). Гематоксилин-пикроиндигокармин. МБИ-6, 10х8 (В. В. Лемещенко): 1 – внутренняя оболочка; 2 – средняя оболочка; 3 – внешняя оболочка; 4 – продольные неисчерченные миоциты.
У животных суточного возраста среди всех афферентных кровеносных сосудов наиболее толстую стенку имеет пупочная вена (рис. 95). В ней преобладает средняя оболочка, сформированная циркулярным слоем неисчерченных миоцитов со значительным количеством эластических волокон и мембран. Во второй афферентной магистрали – воротной вене, на фоне тонкой средней оболочки уже растет адвентициальная, которая содержит продольные пучки мышечных клеток (рис. 96). При этом их больше всего у зрелорождающихся животных – телят. У поросят и, особенно, щенков их количество значительно меньше, что по нашему мнению, указывает на более существенное влияние сердечной деятельности на портальную гемодинамику. Следует отметить, что у поросят с низкой живой массой количество мышечных компонентов в стенке воротной вены значительно уменьшается, предопределяя дополнительную нагрузку на сердце при регуляции системного кровообращения.
В стенке коллектора пупочных ворот и интраорганных вен за исключением участков больших печеночных вен, а также в венозном протоке, содержатся лишь циркулярный слой миоцитов. Его выраженность не равнозначна в венах разных участков долей печени, предопределяя разную регуляторную активность определенных сосудистых сегментов.
Большие интраорганные эфферентные магистрали и коллектор, в брюшной полости при тонкой средней оболочке, имеют развитую адвентициальную оболочку с продольными пучками миоцитов. Такое ее строение указывает на активную регуляцию эфферентного печеночного кровотока сосудистыми факторами. В грудной полости стенка каудально полой вены (коллектора) бедна мышечными компонентами и она выполняет функцию транзитной магистрали кровотока к сердцу.
С возрастом животных, на фоне облитерации пупочной вены и венозного протока в стенке афферентных и эферентных венозных магистралей печени животных увеличивается количество мышечных и волокнистых компонентов их стенки, что предопределенно, возможно, ростом интенсивности кровотока через орган.
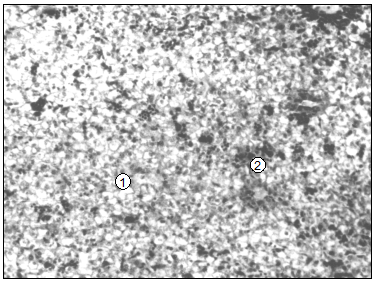
Микроциркуляторное русло печени суточных животных состоит из стромальной и паренхиматозной систем (рис. 97, 98). В стромальную- кровь притекает по артериальным звеньям-артериолам и прекапилярам. Ее капиллярные сетки соматического типа размещены, в основном, вокруг желчных протоков и больших афферентных вен. Из ных оттекает кровь к портальным посткапилярам и венулам, которые являются терминалями междольковых афферентных вен. С возрастом животных эта система почти не изменяется.
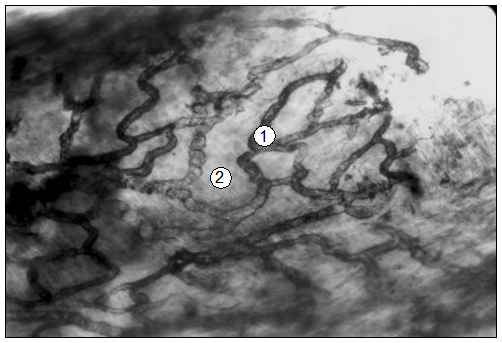
Рис. 97. Гистотопограма печени поросенка (сутки). Инъекция раствора азоткислого серебра. МБИ-6, 10х9 (В. В. Лемещенко): 1 – стенка афферентной вены; 2 – стромальные капилляры.
В паренхиматозную микроциркуляторную систему кровь направляется, как артериальными звеньями (в меньшем количестве), так и венозными прекапилярами портального происхождения. Синусоиды печени у суточных животных не имеют определенной ориентации. Лишь у телят они приобретают радиальное размещение вокруг дренажных звеньев кавальной системы, которые состоят из венозных посткапиляров и посткапилярных венул. Следует отметить, что последние являются терминалями печеночных вен и в литературе их определяют, как центральные вены. Однако, архитектоника, незначительный поперечник и тонкая стенка (в посткапилярах лишь из эндотелия), по нашему мнению, указывает на их принадлежность к микроциркуляторному, а не венозному руслу, которое предопределяет особенности гемодинамики в ных. Такая структура паренхиматозной микроциркуляции указывает на незначительную пропускную способность “чудесной сети” новорожденных животных, которая лишь несколько возрастает в начале формирования структурно функциональных единиц органа.
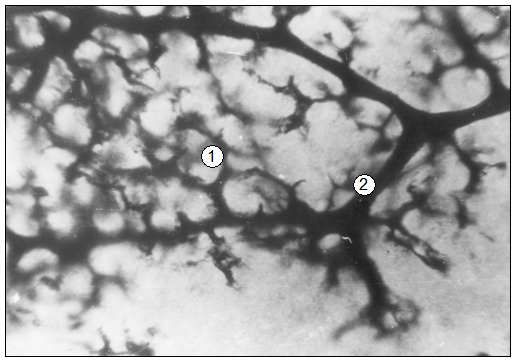
Рис. 98. Просветленный препарат печени суточного щенка. Инъекция туш-желатиновим гелем. МБИ-6,10Х9: 1 – синусоиды; 2 – венозные пост капилляры.
Морфологические данные, относительно активного участия печени и ее вен в регуляции интенсивности кровотока к сердцу, подтверждаются прижизненными исследованиями этих структур с помощью ультразвуковых исследований.
В настоящее время ультразвуковые исследования структур организма животных еще не находят широкого применения в морфологической практике. В ветеринарной медицине их применяют преимущественно с диагностической целью при заболеваниях мелких домашных животных, а также для контроля процессов беременности продуктивных животных [319, 320].
Однако, ультразвуковые исследования кровеносных сосудов у взрослых людей свидетельствуют, не только об их достаточной визуализации, но и выясняют динамику параметров в зависимости от экстракардиальных факторов. Да, нижняя полая вена при дыхании здорового человека способна изменять свой поперечник до 50%, а при пробе Вальсальви (формируемый выдох с закрытым ртом и носом) как нижняя полая, так и печеночные вены расширяются [172, 175].
Наши прижизненные исследования печени и ее вен у новорожденных животных проведены с помощью ультразвукового сканера в В-режиме реального времени, свидетельствуют о постоянстве их параметров в зависимости от вдоха и выдоха. Да, объем печени у телят и поросят до 20-суточного возраста является динамическим, что подтверждается изменением относительной площади и яркости (в условных единицах (УО) Adobe Photoshope, v.6) ее изображения на мониторе ультразвукового прибора (табл. 109, 110).
Таблица 109
Динамика относительной площади и экзогенности печени 1-, 10- и 20-суточных телят (В. В. Лемещенко)
|
Возраст Суток |
Относительная площадь % |
Экзогенность (УО) |
||
|
Вдох |
Выдох |
Вдох |
Выдох |
|
|
1 |
7,60±2,54 |
8,14±1,93 |
94,45±3,61 |
93,06±4,37 |
|
V,% |
57,89 |
41,03 |
6,63 |
8,13 |
|
10 |
10,70±0,74 |
11,50±0,89 |
100,08±6,50 |
98,21±8,11 |
|
V,% |
11,96 |
13,39 |
11,24 |
14,30 |
|
20 |
11,77±3,08 |
14,06±3,23 |
120,00±3,31 |
117,42±6,24 |
|
V,% |
45,28 |
39,76 |
4,78 |
9,20 |
Таблица 110
Динамика относительной площади и экзогенности печени 1-, 10- и 20-суточных поросят (В. В. Лемещенко)
|
Возраст Суток |
Относительная площадь % |
Экзогенность (УО) |
||
|
Вдох |
Выдох |
Вдох |
Выдох |
|
|
1 |
10,16±1,18 |
11,85±0,90 |
57,69±3,73 |
53,00±3,30 |
|
V,% |
23,23 |
15,16 |
12,95 |
12,47 |
|
10 |
9,04±2,44 |
13,01±1,62 |
62,87±1,75 |
56,06±2,66 |
|
V,% |
46,68 |
21,60 |
4,81 |
8,22 |
|
20 |
17,81±2,10 |
18,76±2,34 |
45,90±7,01 |
40,49±7,46 |
|
V,% |
20,44 |
21,64 |
26,43 |
31,33 |
При вдохе у телят и поросят к 20-суточному возрасту уменьшается относительная площадь изображения печени на ультразвуковом секторе при увеличении его яркости и, соответственно, экзогенности. Такая закономерность динамики экзоморфологии печени доказывает увеличение объема органа при его кровенаполнении.
Сканирование екстраорганных афферентных и эфферентных магистралей печени телят к 20-суточному возрасту также доказывает непостоянство их параметров от вдоха и выдоха животных (табл. 111). При вдохе поперечник пупочной вены несколько возрастает, а воротной и каудальной полой наоборот – уменьшается (рис. 99, 100).
Таблица 111
Динамика поперечника вен на сонограмме печени 1-, 10-ые 20-суточных телят. (В. В. Лемещенко)
|
Возраст животных Суток |
Пупочная вена |
Воротная вена |
Каудальна полая вена |
|||
|
Вдох |
Выдох |
Вдох |
Выдох |
Вдох |
Выдох |
|
|
1 |
1,40±0,14 |
1,35±0,16 |
1,68±0,17 |
1,77±0,07 |
1,63±0,12 |
2,11±0,39 |
|
V,% |
14,28 |
18,25 |
14,88 |
5,65 |
11,04 |
26,54 |
|
10 |
1,40±0,43 |
1,32±0,28 |
1,60±0,21 |
1,63±0,05 |
1,72±0,21 |
2,10±0,25 |
|
V,% |
43,57 |
30,30 |
14,48 |
10,19 |
7,60 |
10,00 |
|
20 |
1,53±0,39 |
1,51±0,20 |
1,70±0,12 |
1,77±0,16 |
2,08± |
2,30±0,25 |
|
V,% |
35,95 |
18,48 |
10,00 |
12,99 |
11,06 |
15,22 |
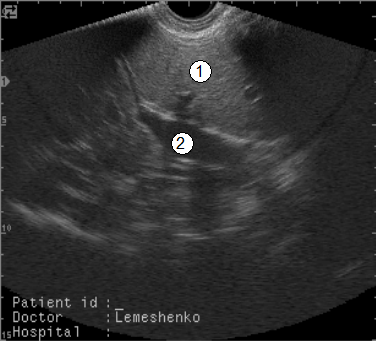
Рис. 99. Сонограма печени суточного теленка (вдох) (В. В. Лемещенко): 1 – печень; 2 – коллектор пупочных ворот.
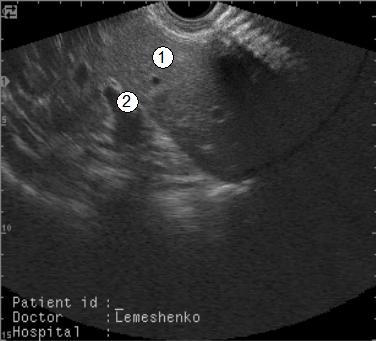
Рис. 100. Сонограма печени суточного теленка (вдох) (В. В. Лемещенко): 1 – печень; 2 – коллектор пупочных ворот.
Следовательно, у новорожденных животных, на фоне адаптогенеза почти всех аппаратов и систем органов к среде дефинитивного существования, происходит глубокое изменение функций печени как органа. В пренатальный период после затухания процессов универсального гемопоэза печень еще не полностью выполняет присущие ей дефинитивные функции пищеварения и дезинтоксикации. В то же время в ней активно протекают процессы дифференциации внутренней структуры с активным накоплением энергетических биохимических веществ, в частности гликогена [20, 34, 55, 296]. При этом в ней, уже из ранних стадий пренатального онтогенеза определяется развитая экстра - и интраорганная кровеносная система, морфогенетически и функционально связанная с сердцем.
По нашему мнению, печень является не только “метаболическим мозгом” организма, но и “гемодинамическим”, особенно в пренатальный и неонатальный периоды онтогенеза. После рождения животных процессы дифференциации в печени несколько затухают. Ее энергетические вещества, которые тратятся во время родов восстанавливаются, однако процессы пищеварения и дезинтоксикации еще не имеют видовой специфичности. В то же время ее кровеносные сосуды продолжают выполнять не только трофические функции, характерные для органа, но и принимают активное участие в регуляции объема крови, оттекающей к сердцу из органов брюшной полости. При этом, как свидетельствуют данные Василевськой С. И. [42], нарушения структуры и функции печени при хронических ее заболеваниях приводят к тяжелому функциональному состоянию миокарда у людей, состояние которого улучшается после лечения заболеваний печени. Такой факт свидетельствует о тесной функциональной взаимосвязи кровеносных сосудов печени и сердца у млекопитающих. Мы считаем, что нарушение кровообращения в печени неонатальных животных приводит не только к развитию общей интоксикации организма, но и к существенным гемодинамическим расстройствам, которые вызывают дисфункцию миокарда.